Be able to explain why gene expression and chromosome
segregation are fundamentally in conflict, and how the
eukaryotic cell cycle resolves this conflict
Be able to describe how DNA is packaged by histone proteins
into nucleosomes
Understand how histones are replaced by protamines in
spermatogenesis
Be able to apply experimental evidence that demonstrates the
'open' chromatin requirement for transcription
Understand the experiments that demonstrate that
transcriptionally active DNA remains associated with histones
Prokaryotes have small genomes,
usually less than 10 7 bp. They get along just
fine carrying out replication, transcription and translation
simultaneously.
Eukaryotic genomes are huge,
ranging from 10 7
bp to 10 12
bp. As we saw in the
exercise, the eukaryotic nucleus has to pack and unpack a
tremendous quantity of chromatin once every cell cycle. That's
a lot of spaghetti to keep organized! For this reason,
eukaryotes need the elaborate mechanisms of mitosis and
meiosis.
2. Two conflicting requirements of the cell:
a. On one hand - DNA needs to be accessible to
transcriptional and replicative enzymes, so that gene
expression and DNA replication can occur.
b. On the other hand - DNA needs to be packaged
tightly so that it can be easily divided between two daughter
cells during cell division.
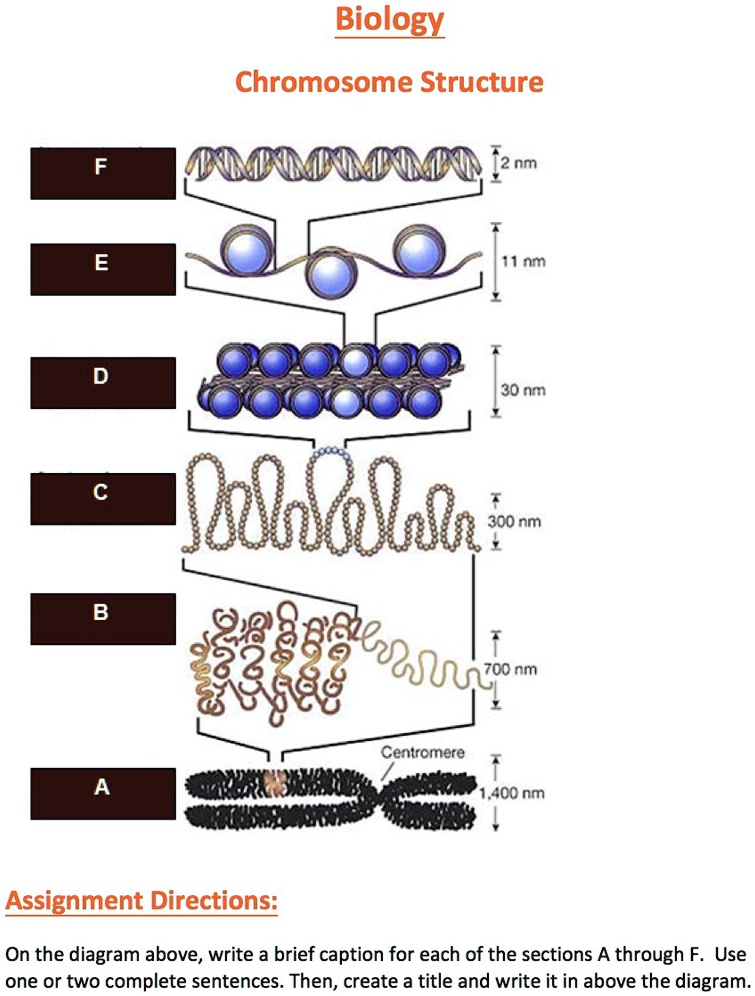
The solution to this conflict is the eukaryotic chromosome.
Chromatin is DNA complexed with proteins. The chromosome
is a single DNA molecule complexed with proteins. It is organized
to allow for a hierarchal packing scheme.
DNA coils around histone octamer particles to form nucleosomes
While there are multiple proteins that complex with DNA, histones
are by far the predominant class of chromatin proteins. Histone
proteins are responsible for helping DNA coil, as well as
regulating transcriptionally "open" and "closed" genes. As they
have an overall positive charge, they attract negatively-charged
DNA. This charge attraction alone is sufficient for nucleosomes to
form. There are five main histone proteins you should know:
Histone Protein
Molecular Weight (kD)
Major Amino Acids
Function
H1
21
Lys++
Linker histone
H2A
13.8
Lys
Core histone
H2B
13.8
Lys
Core histone
H3
15.4
Arg
Core histone
H4
11.4
Arg
Core histone
Nucleosome
structure
Moudreinakis, EN and Arents, G (1993)
Structure of the Histone Octamer Core of the Nucleosome and
Its Potential Interactions with DNA. in Cold Spring
Harbor Symposia on Quantitative Biology Vol. LVIII DNA and
Chromosomes.pp. 273-279. Cold Spring Harbor Press
1993.
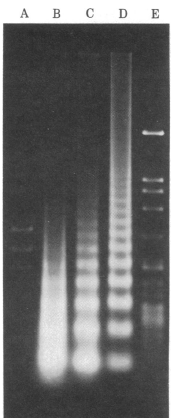
Treatment of nuclei with limiting
amounts of micrococcal nuclease (an endonuclease) typically
results in a "step-ladder" of bands, showing that DNA is cut
into discrete fragments in multiples of ~200bp. If you digest
with higher concentrations of nuclease, the periodicity changes
to 146bp.
Interpretation: something in chromatin is
protecting pieces of DNA, about 146 bp long, from being
digested. Most of the chromosomal DNA is being protected.
MICROCOCCAL NUCLEASE DIGESTION OF
CHROMATIN
[Spiker et al. (1983) PNAS 80:815 Fig. 4] (Note:
this figure compares 3 alternative methods for
chromatin isolation)
Fig. 4. Micrococcal nuclease digestion of wheat embryo
nuclei and chromatin. Nuclei and chromatin substrates were
adjusted to 1µg of DNA/ml and digested with micrococcal
nuclease at 50 units/ml at 37°C. Time course of digestion
experiments were carried out for all samples. The DNA
fragments were then purified and separated
electrophoretically on agarose gels (pH8.1). AHaeIII-digested
phi-X174DNA. B chromatin isolated by the method of
Simon and Becker. A 6min. digest is shown; this extent of
digestion and quantitiy of DNA applied to the gel showas a
nucleosome pattern better than that found under any other
conditions although the faint nucleosome pattern is barely
noticeable against the background of variable length
fragments produced by micrococcal nuclease. C
chromatin isolated by the modified method of Bonner et
al.; a 10 min. digest is shown. D nuclei
isolated as described in the text; a 2-min. digest is
shown. EHaeIII-digested Lambda phage DNA.
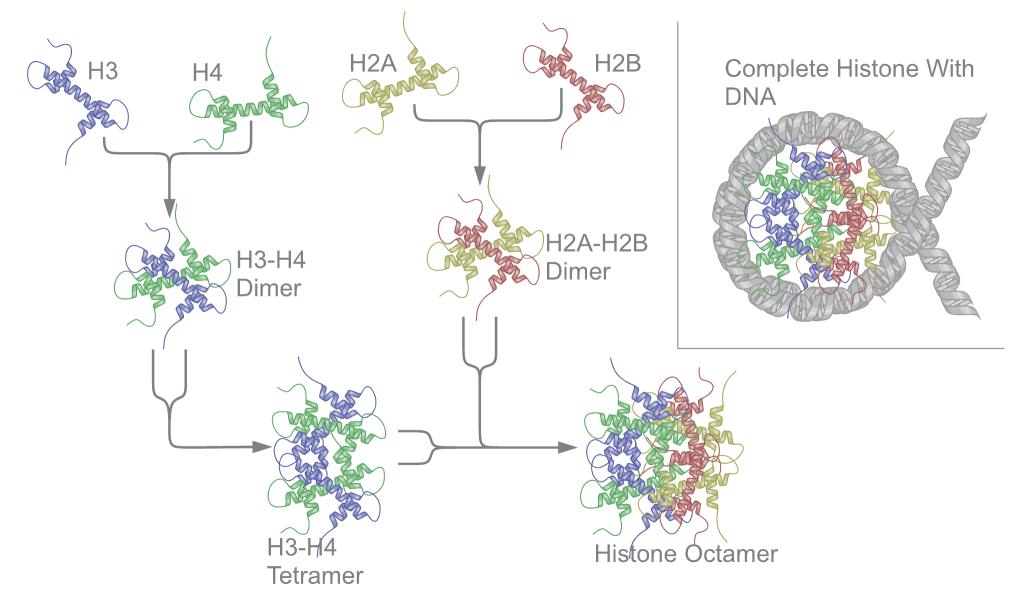
Nucleosome core particle is a histone
octomer, consisting of two molecules each of H2a, H2b, H3 and H4.
Particles have a twofold symmetry.
The histone core particle is an octamer, and
contains 2 copies each of H2A, H2B, H3, and H4. H1 is a
linker histone that has three distinct regions: a basic (+),
random coil at the N-terminus, a central globular region,
and a highly conserved, highly basic C-terminus. DNA coils
around histone octamers to form nucleosomes.
Nucleosomes are the first stage of the coiling process, and
contain the histone octamer and two turns of DNA.
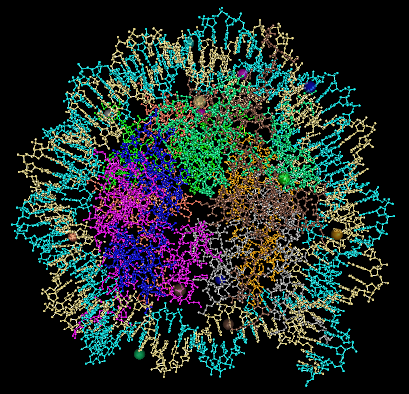
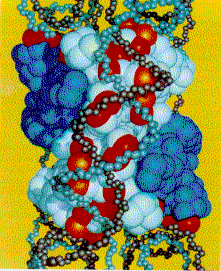
The nucleosome structure includes
the core histone octomer plus 2 turns of DNA. Two rotated
views of the nucleosome are shown in ball-and-stick
representation. The two strands of the DNA double helix
are shown in light blue and cream color. The eight histone
proteins in the nucleosome core are each colored
differently. Wrapped
around these particles is two-turns of a DNA
helix. These are left-handed turns. They take up,
not surprisingly, 146 bp.
These figure were produced from
PDB entry 2CV5, using the NCBI Cn3D viewer.
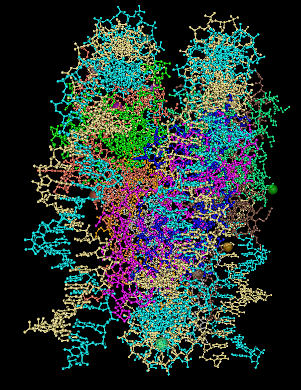
Complementarity between
positive charges on the core protein and the DNA
phosphodiester backbones. The (H3-H4)2
is white and the H2A-H2B dimers are blue. the C-alpha
atoms of lysines and arginines on the cylindrical surface
are indicated by red, and the atoms marking the positions
of the positive end of helix dipoles are indicated by
orange. Atoms of the DNA backbone are "undersized" so as
to allow visualization of the protein surface.
Nucleosomes can form
spontaneously in-vitro
The positive charges of the histone proteins and the
negative charges of the DNA backbone are sufficient to
drive nucleosome formation. In chromatin reconstitution
experiments, histones H2a, H2b, H3 and H4 are added to
DNA, resulting in nucleosome formation. Therefore, the
process of nucleosome formation is solely due to charge.
From:
Moudreinakis, EN and Arents, G (1993) Structure of the
Histone Octamer Core of the Nucleosome and Its
Potential Interactions with DNA. in Cold Spring
Harbor Symposia on Quantitative Biology Vol. LVIII
DNA and Chromosomes.pp. 273-279. Cold Spring
Harbor Press 1993. Fig. 6
Which is the correct model for chromatin fibers: 10 nm, or 30
nm?
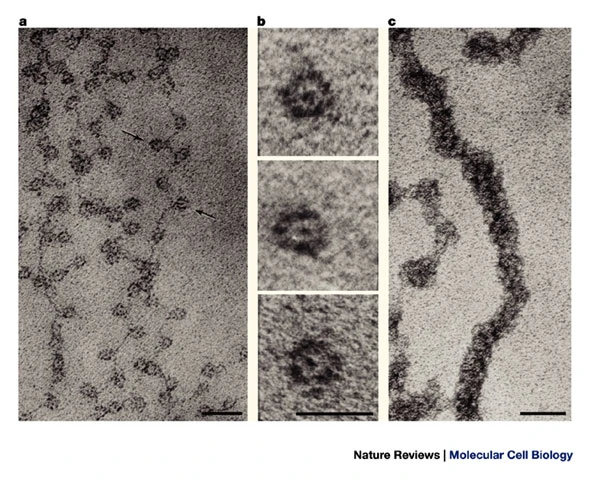
NUCLEOSOMES:
"BEADS
ON A STRING"
If you isolate nuclei
(eg. Drosophila)
and extract chromatin at low ionic strength (<100mM
salt), you see "beads on a string" images in electron
microscopy (a). These beads are nucleosomes (b). An
interpretation of the structure is shown below:
Two turns of DNA are coiled
around each nucleosome, with a short region of naked DNA
linking each nucleosome. Since the width of a nucleosome
is about 10nm, these are called "10nm fibers". (In
contrast, a DNA double helix is 2nm wide.)
If you extract chromatin at
high salt (200-300mM), or re-constitute chromatin by
increasing Na+ or Mg++ conc, you
get a 30nm fiber, or solenoid (c). The
30nm fiber is a coil of nucleosomes made up of 6
nucleosomes per turn.
Image displayed by hyperlink
to https://media.springernature.com/full/springer-static/image/art%3A10.1038%2Fnrm1225/MediaObjects/41580_2003_Article_BFnrm1225_Fig3_HTML.jpg?as=webp
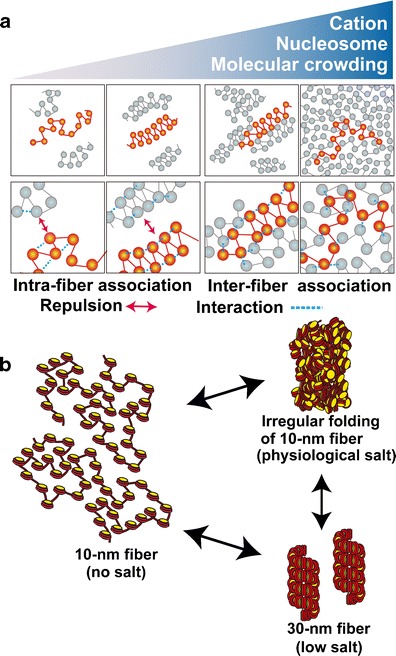
But is the 30 nm fiber
actually there in-vivo?
For many years, it
was thought that the 30 nm solenoid was the default
structure for chromatin in the nucleus and in the mitotic
chromosome. However, that assumption was based on the
in-vitro evidence described above.
There are new reports that indicate that in-vivo, the 30
nm fiber is probably not the predominant structure.
Fussner et al. report that 3D images generated by
combining electron spectroscopic imaging with 3D
tomography find no evidence for 30 nm solenoides in mouse
embryonic fibroblasts. Rather, all chromatin seemed to
be in the 10 nm fiber configuration
Further evidence
from cryo-EM and X-ray scattering, indicate strong
evidence for chromatin at periodicities both 6 nm
and 10 nm, but no evidence for a periodicity of 30 nm, in
both interphase chromatin and mitotic chromosomes.
Tentative
conclusion: The 30 nm fiber is most likely an artifact of
salt concentrations used in chromatin reconstitution
experiments. The current favored model is that the default
state of chromatin in the nucleus, and in the mitotic
chromatin, is the 10 nm fiber.
Alternative model: Irregular folding of 10-nm
fibers.
Since the 30-nm fiber appears not to form at
physiological salt conditions, one model proposes that 10-nm
fibers associate in an unorganized fashion, governed by
hydrophobic and hydrophilic interactions. The result is
irregularly folded bundles of 10-nm fibers of chromatin.
One advantage of this model is that it predicts that
chromatin bundles will be less stable than the
highly-organized 30-nm fibers would be. That means that
bundles will be expected to open and close, to fold and
unfold, giving transcriptional enzymes and DNA binding
proteins better access to genes. Thus, transcription is
easier to initiate on irregularly-folded chromatin, than it
would be in 30-nm fibers.
Open
Access This article is distributed
under the terms of the Creative Commons
Attribution License which permits any use,
distribution, and reproduction in any medium,
provided the original author(s) and the source are
credited.
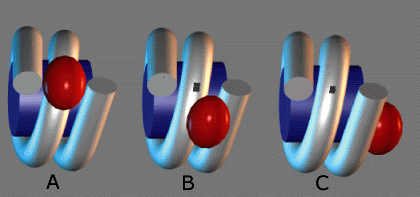
Linker histones are important to coiling of
nucleosomes. Histone 5 is also a linker histone, that is
seen more often in transcriptionally active chromatin.
Histone H1 tends to be seen more in inactive chromatin. The
individual nucleosomes can be linked in three different
ways, as per the diagram at right.
Three arrangements for linker
histones are shown as a) symmetrical, b) bridging, and
c) asymmetrical.
Which linker histone would more likely be present
in
(a) constitutive heterochromatin?
(b) euchromatin?
(c) facultative heterochromatin?
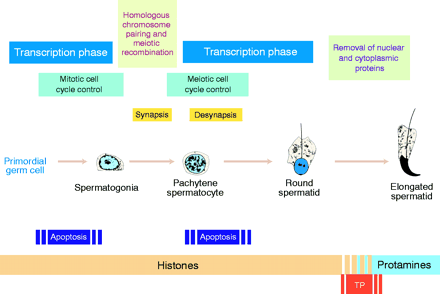
To facilitate tight compaction of DNA in sperm, histones are
replaced by protamines
In mamalian spermatids, the nuclear volume is
about 5% that of the somatic cell nucleus. To achieve this
level of packaging, an arginine- and cysteine-rich class of
proteins called protamines replace
histones. As illustrated in the figure, a round of
transcription occurs in haploid spermatids, producing
copious quantities of protamines, which displace histones in
mature spermatids. In mamalian spermatids, protamines do not
directly displace histones. Mamalian spermatids first
displace histones with basic "transition proteins", which
are in turn displaced by protamines.
From P. Sassone-Corsi (2001) Unique
chromatin remodeling and transcriptional regulation in
spermatogenesis. Science 296: 2176-2178.
For transcription to occur, the chromatin structure of a gene
must be 'open', that is, accessible to the transcriptional
machinery
In this discussion about histone proteins and nucleosomes, you
may have been wondering: if DNA is complexed with these
proteins, won't that make it hard to transcribe DNA?
The enzymes of the eukaryotic transcriptional apparatus are
adapted to the presence of chromatin proteins. This is the price
eukaryotic cells pay for having such large genomes. You can't have
a large genome without having the extra overhead of organizing it,
which is done by the chromatin proteins. Like everything else in
eukaryotic cells, gene expression and replication are very
deliberate and carefully orchestrated processes.
So, for gene expression to occur, the DNA can be complexed with
proteins, but it has to be in a transcriptionally "open"
position. How can we detect this "open" chromatin
structure?
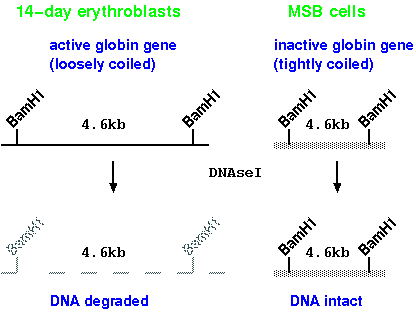
1. General sensitivity to DNase I
We can compare chromatin structure around the
beta-globin gene of chick embryo erythroblasts (precursors
to red blood cells), which do express globin genes, and an
undifferentiated cell line MSB, which does not express
globin genes. In erythroblasts, the DNA of the globin gene
is preferentially sensitive to DNAseI digestion. This
suggests that the structure of the chromatin is looser in
cells expressing globin.
Note: These treatments are done using isolated nuclei,
not naked DNA
Procedure: Isolate nuclei nuclei | Divide into several tubes v |----+----+---+---+--+--| v v v v v v v DNAseI [µg/ml] 0 .01 .05 .1 .5 1 1.5 | Isolate DNA v | Cut with BamH1 v | Electrophoresis v | Blot v |Probe with globin gene v
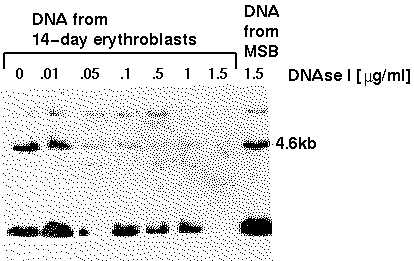
In the
autoradiogram, we see that even at the highest DNAseI
concentration, the 4.6kb fragment is relatively
insensitive to digestion in nuclei from a cell line that
does not express the globin gene. In the erythroblasts,
however, specific degredation of this sequence can
be seen to occur even at the lowest concentration. The
interpretation of this is that the globin gene is in a
more open chromatin configuration, which provides
greater access to DNAseI. This phenomenon is referred to
as general nuclease sensitivity, because the whole
gene appears to be digested. However, more detailed
studies of certain genes can detect specific sites
within transcriptionally active genes that are hypersensitive
to nuclease digestion.
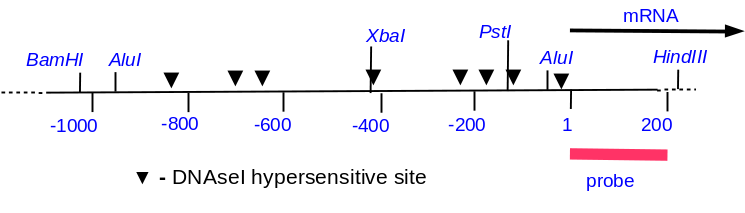
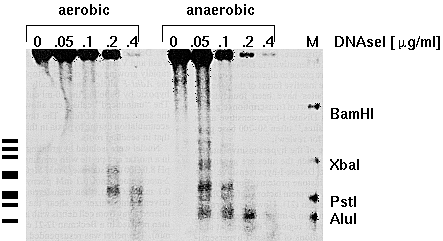
The Adh1 (alcohol dehydrogenase) gene in maize can be induced
(ie. made to produce mRNA) by anaerobic conditions. This helps
the plant detoxify the end products of anaerobic respiration. In
cell culture, anaerobic conditions can be simulated by bubbling
Ar into cell suspensions for 2hr. This system permits the
comparison of chromatin structure of the Adh1 gene between
anaerobic and aerobic conditions. Below is a restriction map of
the region near the Adh1 gene in maize. (Start of transcription
is +1). We can map hypersensitive sites by indirect
end-labeling. Note that in the general sensitivity experiment,
we used a probe that overlapped the entire gene in question.
Since it is possible that many sites in a given gene would be
digested simultaneously by DNAseI, the Bam fragment above was
chopped into pieces too small to detect on an agarose gel. But
with indirect end labeling, we use a short probe that lays
outside the region we wish to examine, from +1 to +210.
M is a
marker lane, with a mixture of DNAs generated by
digesting this gene with either BamH1, Xba1, Pst1
or Alu1, and then mixing all of these DNAs together.Now, when the DNAse-ed
DNA is digested with Hind3, the +1 to +210 probe will only
detect fragments upstream from the Hind3 site. If we do a
limiting digestion with DNAseI, in any given
fragment, only one of several possible hypersensitive
sites will be digested in a given molecule. Thus, we
get a "ladder", in which each rung represents a different
cut site. Note that there are two constitutive
hypersensitive sites, even in uninduced (aerobic) nuclei,
whereas in induced nuclei, we see 8 distinct
hypersensitive sites.
from
Anna-Lisa Paul, Vimla Vasil, Indra K. Vasil, and Robert J.
Ferl. Constitutive and anaerobically induced
DNase-I-hypersensitive sites in the 5′ region of the maize
Adh1 gene. PNAS 84:799-803
Transcriptionally active DNA remains associated with histones,
although nucleosome configuration may be different in active vs.
inactive chromatin
So, we can identify regions that are in a
more transcriptionally "open" position. But what does that
"open" configuration look like with the histones? We know
from electron micrograph data that the 10nm beads on a
string configuration is present during transcription.
What's happening here? Let's start with two hypotheses.
H(0): Transcription cannot occur when DNA is complexed into
nucleosomes
H(1): Transcription can occur when DNA is complexed into
nucleosomes
As a means of detecting whether or not DNA is
bound up with histones, crosslinking mediated by psoralen
was used to covalently crosslink any DNA that was accessible
when cells were treated with UV. The spacer DNA between
nucleosomes is more accessible to crosslinking, resulting
covalent bonds between the two strands. When chromatin is
isolated for Electron Microscopy, the proteins are lost, and
only the DNA remains on the grid. The crosslinked spacer
regions remain as doublestranded helix. The DNA that was
complexed with histones becomes single-stranded, giving
appearance of bubbles on the circular chromosome. Thus, each
bubble tells us which DNA was complexed into nucleosomes.
Since we see bubbles on the entire SV40 genome, and we also
see an RNA transcript coming off of the circle, with
nucleosomes on either side of the transcription site, we can
conclude that nucleosome structure remains intact even
during transcription.
Nucleosomes seem to protect little bubbles of DNA in
transcribing genes from treatment with DNA crosslinking reagents
(eg. 6-methyl psoralen). The bubbles can be shown to be roughly
200bp apart. Only the inter-nucleosome DNA is crosslinked. The
long "tail" on the circle is the growing mRNA transcript.
Nucleosome
structure "opens up" during transcription, in a process
called "chromatin remodeling"
So how does the DNA become accessible to RNA polymerase while
staying complexed with histone proteins? The answer is a process
called chromatin remodeling.
The
precise details of chromatin remodeling are still under
investigation. However, the figure at right illustrates
one model for chromatin remodeling. In (d), D and Tr
domains of the ocatmer "walk" along the double helix by
rotating around the Hinge domain. Thus, while the DNA is
more accessible to the solvent, and hence, more sensitive
to nucleases, the nucleosome never completely dissociates
from the DNA helix.
Image displayed by hypertext link to
https://media.springernature.com/lw685/springer-static/image/art%3A10.1038%2Fnsmb1333/MediaObjects/41594_2007_Article_BFnsmb1333_Fig3_HTML.gif?as=webp
Summary
Eukaryotic genomes are huge, which requires them to have
elaborate mechanisms for replicating and packaging their DNA
Histone proteins are necessary for the hierarchal
organization of DNA
Specialized proteins called protamines replace histones to
achieve high chromatin coiling in gametes
While chromatin remains complexed with histones, it needs to
attain an 'open' configuration in order to be transcribed
 from
Anna-Lisa Paul, Vimla Vasil, Indra K. Vasil, and Robert J.
Ferl. Constitutive and anaerobically induced
DNase-I-hypersensitive sites in the 5′ region of the maize
Adh1 gene. PNAS 84:799-803
from
Anna-Lisa Paul, Vimla Vasil, Indra K. Vasil, and Robert J.
Ferl. Constitutive and anaerobically induced
DNase-I-hypersensitive sites in the 5′ region of the maize
Adh1 gene. PNAS 84:799-803 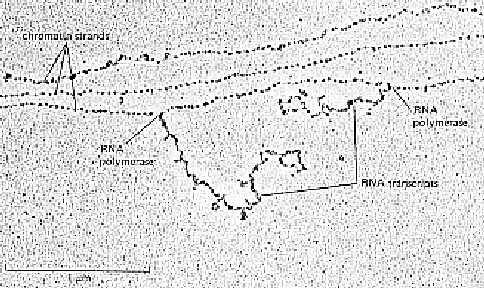


{kind=link}
{kind=link}