
Early prophase I

Late Prophase I
Images from Dept. of Plant Science, University of Manitoba
Flowering plants and fungi go through haploid stages as part of their life cycles. In plants, this is the gametophyte generation. Metabolically-active haploid cells must carry out all fundamental cellular functions. Since only one copy of each gene is present, there is strong selective pressure against deleterious alleles in the haploid state.
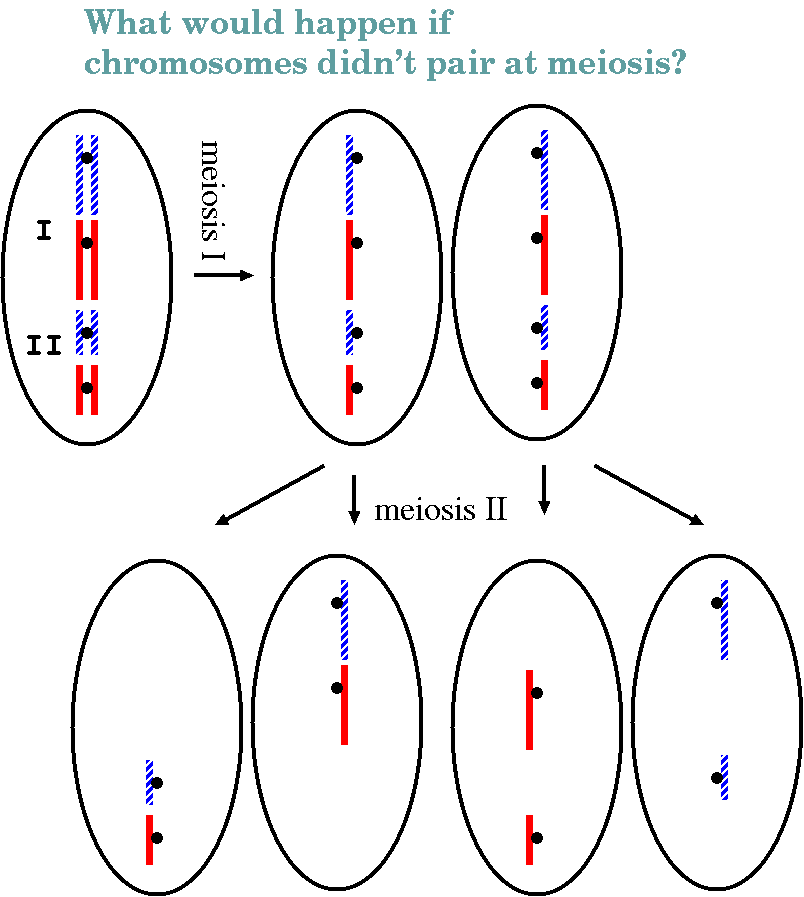
In a nutshell: The diploid cell contains two homologues of each chromosome, one parental and one maternal. These replicate in S phase, resulting in two identical sister chromatids. Prophase I begins with pairing of both homologues, resulting in a tetrad containing 4 homologous chromosomes. Crossing over occurs here. As chromosomes continue to condense, the sites of crossing over become visible as chiasmata. Chromosomes begin to separate as they further condense, pushing chiasmata to the chromosomal termini. As in mitosis, chromosomes are aligned between the centrosomes in metaphase I. One kinetochore forms per chromosome pair, rather than one per chromatid. Consequently, chromatid pairs migrate to opposite poles, such that both chromatids at a given pole are derived from a single parent. In telophase 1, cytokinesis divides the two diploid cells, and in most species, the chromosomes remain condensed and a nucleus does not re-form. In prophase 2, centrosomes divide again, pulling bivalent chromosomes to the center of the cell. Kinetochores divide, and chromosomes migrate to the poles (anaphase 2), followed by telophase 2, in which haploid nuclei form.
Click here for Meiosis summary
Premeiotic interphase can result in an increase in nuclear volume 3 to 4 times that of mitotic nuclei, in preparation for the creation of 4 daughter cells.
Early prophase I |
Late Prophase I |
| Meiotic prophase I in
Rye anthers Images from Dept. of Plant Science, University of Manitoba |
|
Click here for a figure summarizing meiotic prophase I, from Hank Bass' Lab at Florida State University
1. Leptotene (leptonema): chromosomes begin coiling
- During leptotene, there are many independent coiling events
occurring simultaneously along the length of each chromosome.
The chromosomes appear distinct as long slender threads with
many bead-like structures scattered along the length. These
bead-like chromomeres are localized aspects of late
leptotene coiling that later spreads along the length of the
pachytene chromosomes. These are the result of many independent
coiling events happening simultaneously along the entire length
of the chromosome.
Remember that prior to prophase I, DNA replication has already occurred. That means that each thin 'thread' contains two identical chromatids, coiled together. The individual chromatids cannot be resolved at this point.
Chromosomal termini can be seen to be attached to the inner surface of the nuclear membrane at "attachment plaques". Through a "homology searching" mechanism that is still unknown, the ends of homologous chromosomes migrate together on the nuclear membrane, making it possible for synapsis to begin at the termini.
|
2. Zygotene (zygonema): chromosomes pair along their entire length - Think of zippering. Zygotene is characterized by the synapsis of homologous chromosomes. (Remember, each chromosome has two identical chromatids, so at synapsis there are 4 homologous chromatids.) During zygotene, the synaptonemal complex forms between the homologues, apparently aligning them gene by gene. right - Zygotene in pollen mother cell meiosis of Lilium regale. Arrows - sites of synapsisfrom Rickards GK (1965) The Cell Nucleus. Tuatara 13:43. http://nzetc.victoria.ac.nz/tm/scholarly/tei-Bio13Tuat01-t1-body-d5.html#Bio13Tuat01-fig-Bio13Tuat01_059a |
 |
|
The
observation
that telomeres appear to cluster together at the
beginning of meiotic prophase I (ie. leptotene) suggests
that synapsis may be physically coordinated for all
chromosomes at a single location. One simple hypothesis
is that by bringing all chromosome ends together into a
small bundle, it is easier for the homology-searching
mechanism to bring both homologues of each chromosome
together. Click
here
for a Quicktime movie showing a single zygotene nucleus
from maize, in which telomeres hybridized with FITC dye
(green) appear to cluster together in a "bouquet"
formation. Chromosomal DNA is visualized using
DNA-specific DAPI stain (red), and heterochromatic knobs
stained with Rhodamine (white). |
 |
3. Pachytene (pachynema): crossing over occurs - Pachytene is defined as the phase at which chromosome pairing is complete and crossing over occurs.
Structure of synaptonemal complex of the nematode worm, C. elegans.:One role of the Pairing Centers in C. elegans is to promote synapsis, which holds matching chromosomes together during recombination. Fluorescent stains make the proteins of the synaptic "zipper" visible under the microscope
from http://www.lbl.gov/Science-Articles/Archive/LSD-worm-meiosis.html
Meiosis can proceed in the absence of
the synaptanemal complex but there is no recombination. The
synaptanemal complex brings paired chromosomes into stable
alignment during which crossing over can occur.
4. Diplotene (diplonema): pairing is undone
Chromosomes undergo desynapsis, which is accompanied by further condensation of chromatin. Chromatids composing the bivalents begin to separate at one or more points, including the centromeres. The individual chromatids may be visible due to contraction and dissolution of synaptonemal complex. The diplotene bivalent consists of two distinct pairs of chromatids which appear to be attached solely at the chiasmata (singular: chiasma ), the X shaped attachments between the chromosomes. |
 Diplotene in Rye anthers Image from Dept. of Plant Science, University of Manitoba |
The chiasmata are the visible
consequence of recombination events which occurred at pachytene.
As would be expected of crossover products, each chiasma
involves one sister chromatid from one chromosome and one from
the other chromosome.
The number of chiasmata is dependent on the species and the length of the chromosome. The longer the chromosome, the more chiasmata. For example, up to 12 chiasmata per chromosome are seen in Vicia faba.
Terminalization - As the centromeres move apart, the chiasmata slide over the cross-over points along the chromatids that are involved in the exchange toward the distal portions. (The physical process is analogous to pulling apart two pieces of rope that are tangled together.) As they reach the chromosome ends they become arrested there forming terminal chiasmata, locked at the telomeres. The purpose appear to be holding homologues together until metaphase congression is completed. Extreme tension is exerted on the terminal chiasmata, because the chromosomes are being pulled in opposite directions by the spindle fibers, and the terminal chiasmata are all that hold them together.
In animal oocytes, chromosome decondensation can occur at this point, allowing RNA synthesis. Oocytes produce RNA required for production of proteins during the first few rounds of cell division after fertilization. After RNA synthesis is completed, oocytes can halt at diplotene for years before completing meiosis. In animal oocytes, chromosome decondensation can occur at this
point, allowing RNA synthesis. Oocytes produce RNA required for
production of proteins during the first few rounds of cell
division after fertilization. After RNA synthesis is completed,
oocytes can halt at diplotene for years before completing meiosis.
| 5. Diakinesis: chromosomes remain paired only at termini - Chromosome condensation continues, and all four chromatids comprising the tetrad are now distinct. Each bivalent is clearly seen to contain 4 separate chromatids, with each pair of sister chromatids linked at their centromere, while non-sister chromatids that have crossed over are linked by chiasmata. As diakinesis proceeds, the bivalent chromosomes shorten and thicken so that the double nature of each half of the bivalents is undetectable. Rod or ring bivalents are evident. The nuclear membrane disassembles and the bivalents attach themselves by their centromeres to the rapidly formed spindle. |  Diakinesis in Rye anthers Image from Dept. of Plant Science, University of Manitoba |
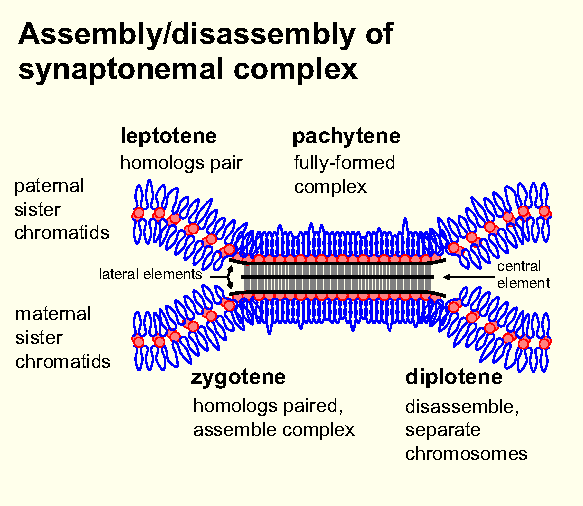
A summary
of the stages of synapsis and desynapsis.
From:
http://users.path.ox.ac.uk/~pcook/students/11mtmebm.html
 |
Attachment
of
spindle fibers from one pole or the other to any given
kinetochore is a random process. Only one spindle can be
attached to each kinetochore. Consequently, bivalents are
arranged at random with respect to which spindle pole is
related to the maternal or paternal centromeres. This is the physical basis for
the genetic concept of random assortment which
determines the meiotic segregation and distribution of
maternal and paternal chromosomes to the two daughter
cells. It forms the basis of Mendel's Law of
independent assortment of non-linked loci. from http://www.wikihow.com/Image:Coin-Toss-1.jpg This file is licensed under Creative Commons Attribution 2.0 license. |
| There
is
a short prometaphase which ends when the bivalent
chromosomes are arranged on the metaphase plate of the
spindle.
The chromosomes are short
and thick. Each bivalent has two kinetochores. The two
sister chromatids are attached to each centromere of the
bivalent. Each of the two centromeres is joined to the
nearer pole by spindle fibres. The two halves of the
bivalent are joined by one or more chiasma and the shape
of the bivalent is determined by the number and
positions of chiasmata in each arm. (That is, we don't
see a perfect 'X' because of the terminal chiasmata.) |
 |
 |
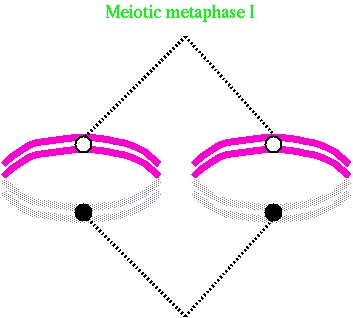 |
In meiosis, the centromeres of the homologous chromosomes are oriented on the long axis of the spindle equidistant from the equator and the terminal chiasmata are located on the equatorial plate.
Equilibrium is established when all bivalent chromosomes have reached equatorial plate until all chromosomes yield to tension of spindle fibres from opposite poles.
This is a good stage for observing chromosome numbers and such conditions of chromosome pairing as univalents, bivalents, trivalents, quadrivalents etc.
During anaphase I, the arms of dyad diverge as the chromatids appear to repel each other. The chiasmata slip off the ends of the chromosomes and the poleward-moving chromatids are bound together at only one point, the centromere. The dyads appear as a double V if chromosome is metacentric or acrocentric and as a single V if telocentric. If no crossover has taken place, the separation is purely reductional. If crossover has taken place, each separating dyad carries part of its homolog, resulting in an equal division of the exchanged chromosomal material to each daughter cell. Chiasmata exchanged at meiosis will redistribute chromosome material from both parents to the daughter cells.
Errors in Anaphase I have serious consequences for the
cell. Any chromosomal connections between the two separating
chromosomes form "bridges" often accompanied by fragments of
chromosomes that lack a centromere (ie. acentric) and therefore do
not move to either pole, lagging on or near the metaphase plate.
Another aberration of anaphase I is nondisjunction - the two
homologs do not separate and/or both go to the same pole.
If no crossover has taken place, the separation is purely reductional. If crossover has taken place, each separating dyad carries part of its homologue, resulting in an equal division of the exchanged chromosomal material to each daughter cell. Chiasmata exchanged at meiosis will redistribute chromosome material from both parents to the daughter cells.
Anaphase I is shorter in duration than Metaphase I. The function of the stage is to evenly distribute the partners of homologous chromosomes to the daughter nuclei and this reduces the number of chromosomes by half. If the somatic chromosome number is 2n= 14, the number is reduced to the gametic chromosome number n=7. Somatic counts at mitosis stated as 2n and a chromosome count in meiosis is stated as n.
| Aguiar-Perecin, Margarida L.R. de, Fluminhan, Antonio, Santos-Serejo, Janay A. dos, Gardingo, José R., Bertão, Mônica R., Decico, Maria Juliana U., Mondin, Mateus. (2000). Heterochromatin of maize chromosomes: structure and genetic effects. Genetics and Molecular Biology, 23(4), 1015-1019. https://dx.doi.org/10.1590/S1415-47572000000400047 | 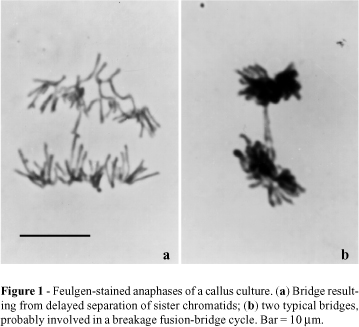 |
This stage is similar to mitotic telophase as the chromosomes assemble at the poles. Nuclear membranes and nucleoli may develop and eventually two daughter nuclei with diploid chromosome number are produced. The chromatids are still widely separated from each other and show no relational coiling. Telophase I in Rye anthers Image from Dept. of Plant Science, University of Manitoba |
 |
Interkinesis is defined as the period between telophase I and
prophase II. During interkinesis, the chromosomes do not
synthesize new DNA and there is no duplication of chromosomes. The
chromosomes are already prepared for the second division each
consisting of two chromatids only held together by the centromere.
Therefore, despiralization, uncoiling and hydration of chromosomes
are not necessary. The length of time between meiosis I and
meiosis II may vary greatly between species.
Prophase II.This second meiotic division is very similar to a mitotic division. Prophase II differs from Prophase I in appearance as the sister chromatids arms are widely separated from each other. This makes the dyad arms look like crosses and the shortness of the partially coiled chromosomes makes it easier to count chromosomes. |
 Prophase II in Rye anthers Prophase II in Rye anthersImage from Dept. of Plant Science, University of Manitoba |
| Metaphase
II |
Anaphase
II |
|
|
 Anaphase II in Rye anthers Image from Dept. of Plant Science, University of Manitoba |
Telophase II |
 Telophase II in Rye anthers Image from Dept. of Plant Science, University of Manitoba |
Evolution
is constantly reinventing the mechanisms of sexual reproduction.
We will only
discuss one example here. In this example, we will detail one of the many
variants of the process of gametogenesis across the higher
eukaryotes, as found in flowering plants.
Microsporogenesis - male meiosis
in flowering plants
The primary function of meiosis in sexually-reproducing species is to produce gametes. This process, called gametogenesis, varies in details among the higher eukaryotes. However, almost all gametogenic processes involve asymmetrical cell division in some way. The examples we've just seen are from gametogenesis of rye anthers, which you might recall is the 'male' gamete in plants. This process produced four similarly sized cells. Certainly they contain different genetic information (that's the whole point of meiosis!), but they are functionally and structurally similar, if not mostly identical. Once meiosis is finished, each haploid cell, called a microspore, develops further into a pollen grain.
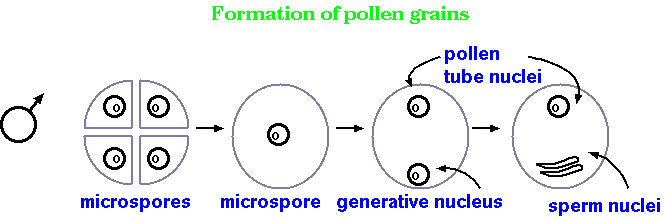
Megasporogenesis - female
meiosis in flowering plants
After going through meiosis, the 4 resulting cells do not
each form an ovum. Instead, three of the cells are designated as
polar bodies, and the primary nucleus undergoes further divisions.
One nucleus from each pole known as polar nuclei migrates to the
middle of the embryo sac and the two nuclei fuse to give rise to a
secondary nucleus or polar fusion nucleus with 2n chromosome
constitution. At the micropylar end of the embryo sac, one nucleus
differentiates into an egg cell and the remaining nuclei become
synergids, part of the egg apparatus. The other three nuclei are
called the antipodal nuclei. During pollination in higher plants,
the tube nucleus of the pollen grain directs the growth of the
pollen tube down the style, through the micropyle and into the
nucellus. The two sperm are released into the nuclear sac. The end
result, after fertilization, is an ovum with 2N chromosomes,
surrounded by a 3N endosperm.
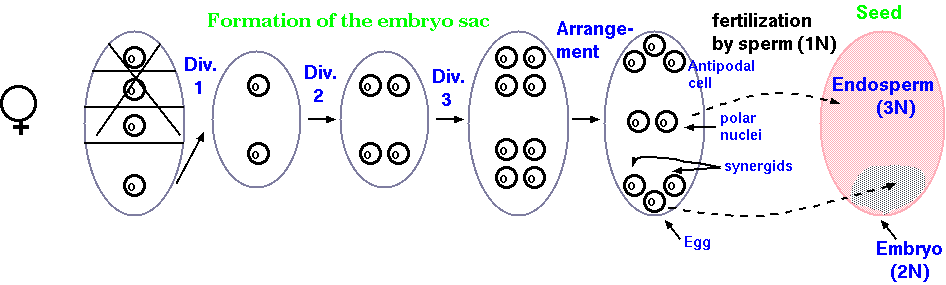
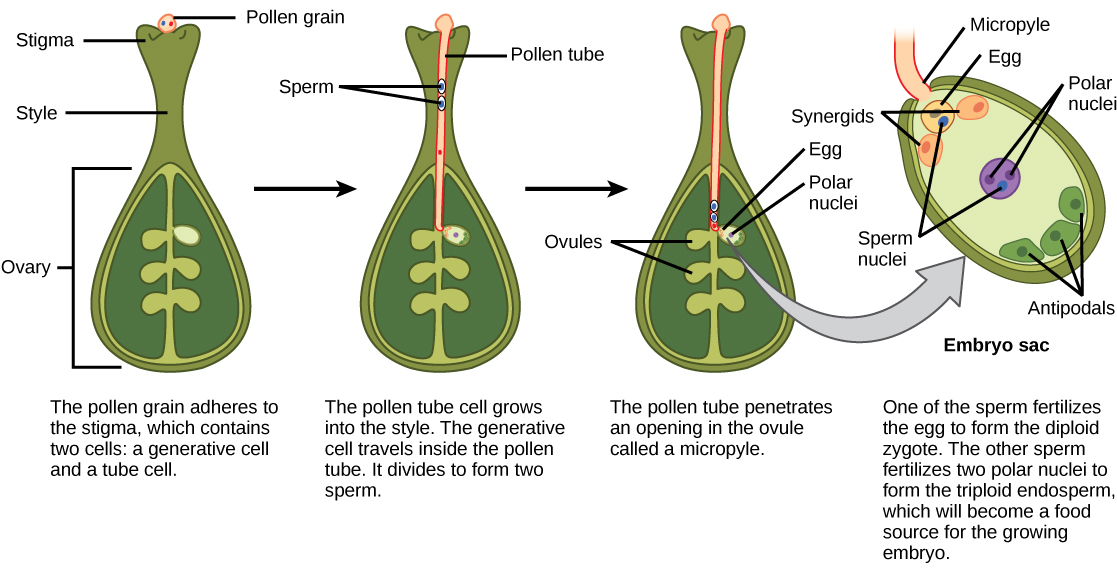
Image displayed from: http://philschatz.com/biology-book/resources/Figure_32_02_07.jpg
In humans, the process is similar in some ways and different in others. Instead of microsporogenesis and megasporogenesis, we have spermatogenesis and oogenesis, respectively. The relative sizes of the gametes are similar to plants: human ova are approximately 10,000 times larger than human sperm. In developing those gametes, we see again the theme of asymmetrical cell division.
Actually, spermatogenesis, like microsporogenesis, produces four
functional gametes after meiosis. Oogenesis is where we see
asymmetrical cell division. Similar to megasporogenesis, we also
see the formation of polar bodies and only one ovum per meiosis.
Most of the cytoplasm stays with the ovum, with only a small
amount of cytoplasm going to the polar bodies. Although the fate
of the polar bodies is generally to disintegrate through
apoptosis, in some species the polar bodies may play other roles.
 Displayed from https://old-ib.bioninja.com.au/higher-level/topic-11-animal-physiology/114-sexual-reproduction/gametogenesis.html
Displayed from https://old-ib.bioninja.com.au/higher-level/topic-11-animal-physiology/114-sexual-reproduction/gametogenesis.html

{kind=link}
{kind=link}