Be able to define and provide and example of a deletion in
genetics, and understand the effects
Be able to define and provide an example of a duplication in
genetics, and understand the effects
Be able to explain how mechanisms (such as unequal crossing
over) can create changes in gross chromosomal structure
Understand the effects of transposons, and the requirements
for transposition
Changes to chromosome structure are often from the cell's
machinery repairing broken chromosomes
Sometimes, usually when the cell is performing regular
activities, DNA will experience a break. Most of the time, the
cell is able to repair the DNA without any problems, and go about
its business. However, some of the time the repair process is
completed erroneously, and this gives rise to some kind of
mutation. It's hard to talk about each of these classes of
mutations separately because all are the result of the cell's
machinery. There are four main classes:
Deletions (deficiencies)
Duplications
Inversions
Translocations
What's important to know is that often, a single event produces
more than one effect. For example, duplications and deletions
often occur together.
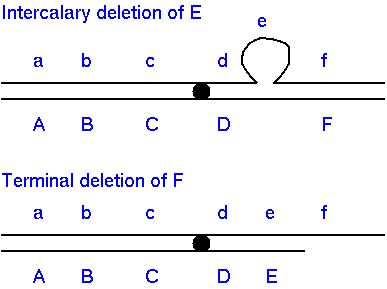
Deletions (deficiencies are the irreversible loss of loci
The loss of a
chromosome segment from a normal chromosome is called a
deletion or deficiency (Df). Deletions can be either
intercalary or terminal.
Terminal
deletions : deletion from the end of a chromosome,
not including the centromere.
Intercalary
deletions - the loss of intercalary or
interstitial segments requires two chromosome breaks and
rejoining of the flanking chromosomal fragments.
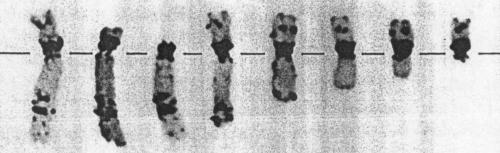
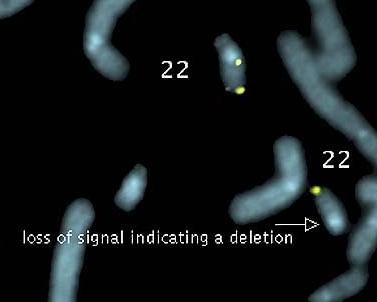
The image below shows a depiction of chromosome 22 from individuals
with DiGeorge Syndrome.
DiGeorge Syndrome is
characterized by cardiac and immune system defects and mental
retardation due to a (cytologically) small deletion on
chromosome 22. Notice that in each pair, one
chromosome is noticeably shorter than the other. This is due to the
deletion of a bright band in the long arm of the chromosome to the
right, near the centromere.
FISH with a
chromosome 22-specific probe shows that two loci
hybridize in a complete copy of chromosome 22, but only
one locus lights up in a DiGeorge chromosome 22.
Consequences of deletions
The size of the deletion can vary from the loss of a nucleotide
to loss of large chromosome segments, and effects become more
serious the more genetic material that is lost. The genetic proof
which can distinguish deletions from point mutations is the
failure to mutate back to the original form. If the deletion
contains a centromere, the resultant acentric fragment will be
lost.
Duplications create additional copies of genes, which can be
advantageous
A duplication is an extra piece of chromosome segment on the
same homologous chromosome (intra-chromosomal) or transposed to
one of a nonhomologous chromosomes in the genome
(inter-chromosomal). The size of the doubled segment can vary
considerably. Duplications are generally more readily tolerated
than deletions, often because the cell still has the minimum
genetic information.
Types
of duplicationsDeletions
and duplications are identified both cytologically and molecularly
Depending on the size of the structural change, duplications and
deletions may be visible as an unpaired segment at pachytene or by
pairing patterns on banded chromosomes. As we have seen above, the
deleted segment of chromosome 22 in DiGeorge Syndrome is large
enough to be detected under a microscope.
In plants, transmission genetics can also be
used to detect changes. The frequency of transmission is variable
depending on the viability of the megaspores containing the
duplication or deletion. (Remember that megaspores and pollen are
haploid, so a deletion means that some genes aren't present at all
in gametes carrying the deletion.) The frequency of transmission
through the pollen is very low. This is either due to the
nonviability of the pollen containing the duplication or deletion
or the failure to compete with the normal pollen in fertilization.
This is another example of the value of the haploid gametophyte
generation in flowering plants. In haploids, deletion of critical
genes can greatly reduce viability of gametes. In animals this is
of little consequence because little gene expression occurs in
gametes, particularly sperm. In maize, dup-del kernels borne on
dup-del plants often make up less than 1/3 of the kernels as the
dup-del gametes are transmitted almost exclusively through the
female- egg cell.
Many
chromosomal changes can be detected using FISH.
As discussed previously,
interphase chromosomes tend to remain in discrete "chromosome
territories", as opposed to being dispersed throughtout the entire
nucleus. Use of two or more locus-specific probes can detect
deletions, duplications, inversion or translocations.
Lengauer C, Kinzler KW and Vogelstein B (1998) Nature
396: 643-649.
c)
Loss of chromosomes 3 (red arrows) and chrom. 12
(yellow arrows) in colorectal cancer cells.
Interphase chromosomes were hybridized with centromeric
probes for chrom. 3 (red spots) and chrom. 12 (yellow
spots). Normal diploid nuclei contain two red and two
yellow spots each.
d) Chromosome translocation. Metaphase plate from
neuroblastoma cells was hybridized with
chromosome-painting probes specific for chromosome 1 (red)
and chromosome 17 (yellow) revealing a t(1;17)
translocation. (Note that chromosome 17 is small, compared
to chrom. 1)
Gross chromosomal structural change can be the result of many
mechanisms
Unequal crossing over creates duplication and deletion
combinations
If unequal crossing over occurs between two homologous
chromosomes, one chromosome will gain a duplication and
the other chromosome will contain a deletion. This is
sometimes referred to in the literature as a dup/del
for short.
One example is the Bar locus in
Drosophila a duplication of 16A segment from region
16A1 to 16A6 of chromosome X which contains 5 bands.
Origin of
Bar-double by unequal crossing over in the Bar-locus
of the salivary gland X chromosome of Drosophila
melanogaster (Redrawn from Morgan et al., 1935. Figure 12. Cold
Spring Harbor Press, New York.)
If this region
is duplicated (Bar mutation), the facets in the
eye are reduced in number (homozygous Bar
average number of facets is 68) which narrows the normal
round eye to a bar-shaped eye. If unequal crossing over
occurs between two chromosomes with the Bar duplication,
a Bar double is produced which in heterozygous
condition reduces the eye facets to 45. Gene expression
is stronger when the duplicated genes are in tandem than
when they are on separate chromosomes (position effect).
Fig. 12.6. Illustration of
the different sizes of compound eyes of the female Drosophila
melanogaster as caused by the varying numbers of facets.
The size of the eye is influenced by the position effect. (From
Kin, 1965. Redrawn from Oxford University Press, New York).
When chromosomal material from another organism is incorporated,
it is called an alien chromosome
The presence of alien chromosomes in a genome increases the
incidence of chromosome structural changes. This has been observed
in wheat (Triticum) lines in which chromosomes from Aegilops
were maintained (monosomic). The mechanism appears to be increased
incidence of chromosome breakage. There is support for this in the
higher number of deletions occurring in the chromosomes with the
higher amount of heterochromatin. Endo and colleagues produced
numerous deletions by introducing Aegilops cylindrica
chromosomes into hexaploid wheat cv. Chinese Spring (see figure
below). These deletions have been used for mapping genes on the
wheat chromosomes. The number of deletions is greatest in the
B-genome and these chromosomes are more heterochromatic than
either the A or D genome chromosomes. A series of deletions were
detected using Giemsa C-banding technique in chromosome 5B.
Observations of deletions and translocations were made in almost
half the progeny of a wheat line in which a single species have
been added through crossing and selection.
Figure 6.2 A series of deletions in
chromosome 5B (normal chromosome 5B extreme left) detected
by Giemsa C banding technique. The horizontal line
represents the kinetochore. (From Endo, T.R. 1990. Jpn. J.
Genet. 65:135-152.)
The ability of wheat to tolerate such drastic chromosomal
deletions is probably due to the fact that it is hexaploid. Most
genes are therefore present on three homologous chromosome pairs.
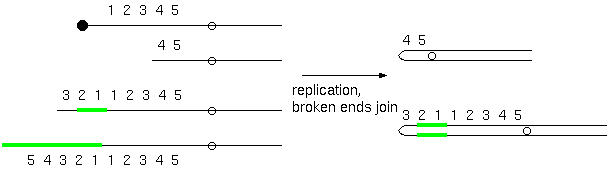
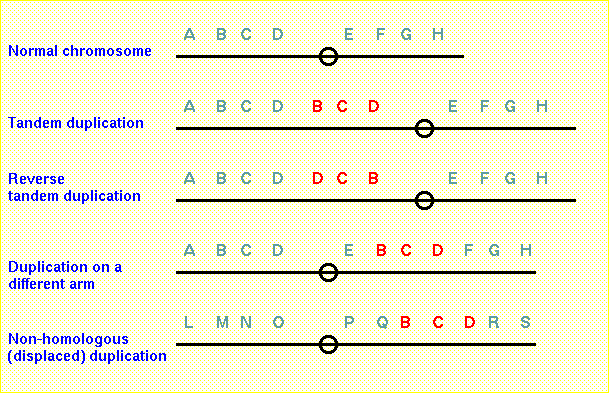
Double stranded breaks can also result in duplications and
deletions
Just
about
any mechanism that can generate double-stranded breaks
could result in duplications or deletions. Tandem, reverse
tandem and duplications in a different arm are
intrachromosomal while displaced duplications are
interchromosomal.
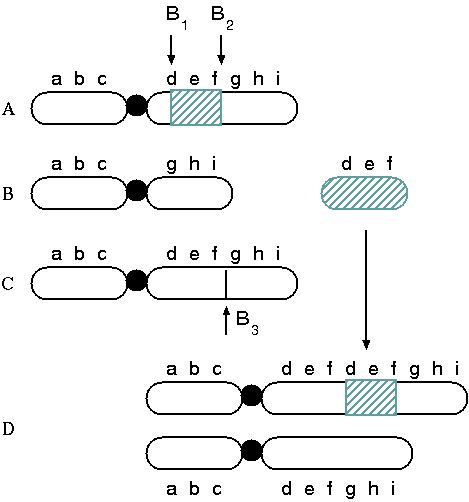
Diagram of a tandem chromosome duplication. (A) The
first two breakpoints (B1 and B2)
in the normal chromosome result in (B) a deleted centric
chromosome (abc.ghi) (C) and an acentric fragment (def).
If the third break occurs in a homologous chromosome (B3
), the acentric fragment (def) could insert into the
partner chromosome resulting in (D) a tandem
duplication.
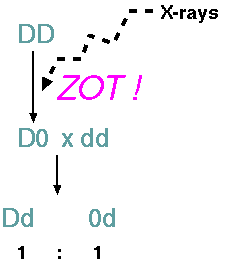
Example:
Irradiation
Induced
deletions can be generated by irradiating pollen. The
pollen is then applied to the stigmas of plants carrying
recessive alleles at loci which may be in the region of
the induced deficiency. In the progeny a few plants will
show the recessive phenotype. (Normally, all F1's should
have the dominant phenotype.) This phenotype is an
indication that the recessive allele is carried in
hemizygous condition due to a deletion in the
corresponding loci on the chromosome from the irradiated
pollen. Genetically, deletions are indistinguishable
from point mutations. Further cytological analysis is
required to verify a deficiency.
If these chromosomes
are examined cytologically at pachytene, one region of
chromosome may show a loop if the deficiency is long enough and
located in an interstitial region. A terminal deficiency results
in an unpaired end region. X-ray induced deletions occur in
either heterochromatin or euchromatin.
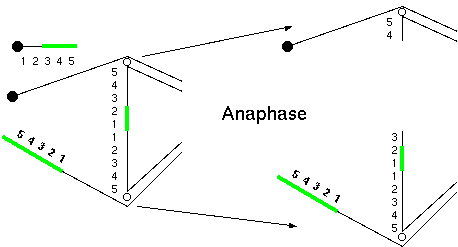
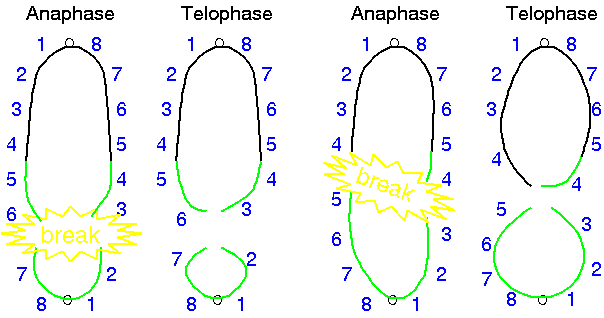
Breakage-fusion bridge cycles are the result of inversion or
translocation in earlier generations
Duplications and deletions can arise in the progeny of inversion
or translocation heterozygotes in which crossing over produces
dicentric bridges at meiosis.
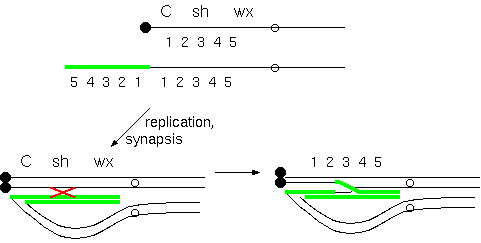
Diagram of a reverse
tandem duplication in the short arm of
chromosome 9 of maize initiating a breakage-fusion-bridge
cycle. A terminal inverted tandem repeat (54321) can
recombine with the homologous region (12345) if the
chromosome with the repeat folds back to allow homologous
regions to pair. In this example, only two chromosomes
participate in crossing over. Therefore, it is only
necessary to pay attention to the two recombinant
chromosomes,since the non-recombinants will be unchanged.
A crossover between 2 and 3 yields one acentric chromosome
containing the "345" region from the distal copy of the
repeat.
At
anaphase, the other recombinant chromosomes will be
dicentric, and will break at anaphase. In this figure, the
break is shown to occur between 3 and 4.
In the next cell cycle, the
broken ends of each mutant chromosome will be joined,
resulting in dicentric chromosomes that will break again
at the next anaphase. (Modified from McClintock, 1941).
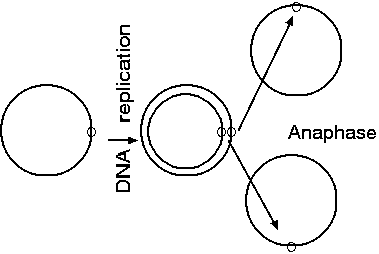
Ring chromosomes are formed from the fusion of broken
chromosome ends
The cellular mechanisms for repairing
double-stranded breaks will sometimes cause chromosomes with
broken ends to fuse, forming ring chromosomes.
Ring chromosomes have been found in several plant species:
maize, tobacco, Antirrhinum, Petunia,
barley, as well as Drosophila and Homo
sapiens. McClintock in papers from 1931 - 1941
studied the behaviour of ring chromosomes in mitosis and
meiosis. Ring chromosomes are not stable
during cell division and are often eliminated. McClintock
developed the breakage-fusion-bridge cycle hypothesis to
explain the changes in chromosome size which occurred as a
result of the ring chromosomes. If the ring reproduces
itself in interphase with no sister strand cross-over in
prophase, the ring chromatids can separate in anaphase
without difficulty resulting in two equal sized
ring-chromosomes, the same size as the original. If sister
chromatid exchange occurs, a ring of twice the size is
produced with two centromeres
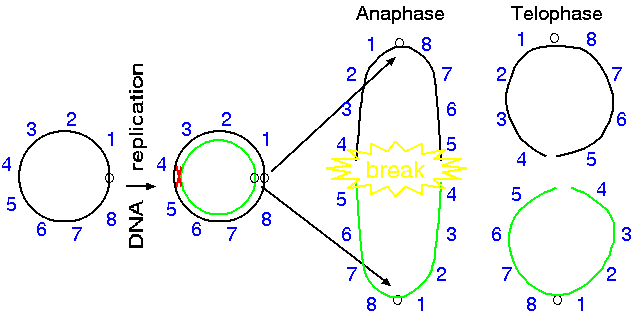
Normally, ring chromosomes can segregate evenly during anaphase.
However, if a crossover event occurs, the two daughter chromosomes
effectively act as a single, circular, dicentric chromosome. The
two centromeres move to opposite poles in anaphase and form an
anaphase double bridge. As the centromeres get pulled to opposite
poles, the chromosomes break at two places. During telophase,
chromosome ends tend to recircularize, in the absence of telomeric
sequences. If the break occus as shown below, balanced chromosomes
will result, with neither deletions nor insertions.
Breaks are possible at different points along the ring
chromosome. If breaks occur asymmetrically, deletions and
duplications can result.
Transposable
elements move from one chromosomal location to another
Structure
Transposition is the
movement of a region of chromosomal segment from one location to
another. DNA is excised from one location by an enzyme called a
transposase, and inserted elsewhere, either on the same chromosome
or a different chromosome. The DNA that is transposed is
referred to as a transposon. Transposons are typically very
small pieces of DNA, ranging from less than 1 kb to several kb.
Transposases
can
cleave and rejoin double-stranded DNA at specific
recognition sequences. When these sequences occur in an
inverted repeat orientation, a transposase can remove the
target sequence from its original location, and insert it
elsewhere. The insertion is essentially a reversal of the
excision.
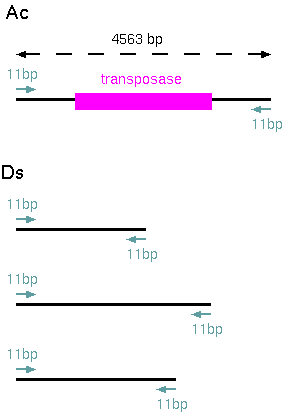
The Ac element
of maize is illustrated at right. It contains a
transposase gene flanked by 11bp Terminal Inverted
Repeats that are recognized by the transposase. Thus,
the Ac element can cause its own transposition.
5' end
(C/TAGGGATGAAA)
3' end
(TTTCATCCCTA)
Other
transposeable elements, such as Ds in maize, are flanked
by the required 11bp repeats, but do not encode a
transposase. Therefore, Ac must be present at another
locus to produce the transposase. In maize lines with Ds
but no Ac, Ds elements do not transpose. When an
Ac-bearing line is crossed with a Ds line, the Ds
elements can transpose. This is referred to as
"mobilization of Ds elements".
Du C (2011) The complete Ac/Ds transposon
family of maize. BMC Genomics 12:588. DOI: [10.1186/1471-2164-12-588]
Discovery of transposition occurred early in the 20th century,
in maize
Ds was first discovered by Barbara McClintock because one of the
Ds elements on chromosome 9 contained a sequence that led to
frequent chromosome breaks. These breaks always occurred at the
same place on chromosome 9, leading to a breakage-fusion-bridge
cycle as described above. Surprisingly, when McClintock crossed a
Ds line with other maize lines, Ds was mobilized to other loci,
causing breaks at those loci.
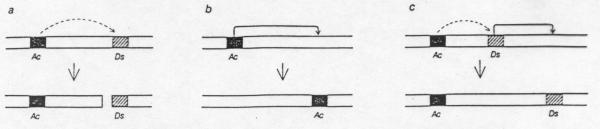
Activator (Ac) locus, was named by Barbara
McClintock for its ability to activate chromosome breakage at
another locus; Dissociation, or Ds (a). The two
loci are shown here on the same chromosome, but they can be on
different chromosomes. Ac is able to promote its own transposition
(b), or that of Ds (c) to another site either on the same
chromosome or on a different one. Ds cannot move unless Ac is
present in the same cell. Ac is an autonomous transposable element
and Ds is a nonautonomous element of the same family.
McClintock's Activator-Dissociator system in maize is a possible
origin of chromosome structural changes. This is a two gene
system: the Ac locus and the Ds locus. When both loci are present,
chromosome breakage increases leading to an increase in chromosome
structural changes: deletions, duplications, inversions,
translocations and ring chromosomes. Ac and Ds are visualised as
blocks of heterochromatin that move by transposition between
different sites in the chromosomes. Ac does not have a mutating
effect alone but promotes the movement of Ds (Ac= activator).
Transposon insertions can cause reversible mutations
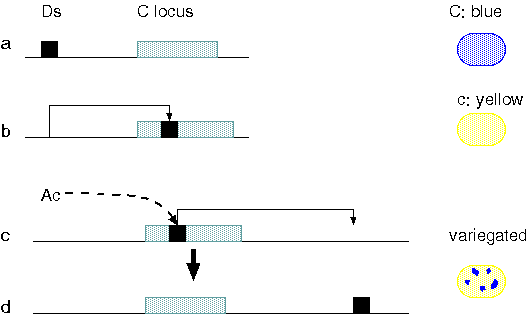
Example: The C locus produces an anthocyanin pigment, resulting
in colored aleurone ie. wild-type. Insertion of a transposon into
C blocks pigment production, resulting in the colorless (c)
phenotype. During seed development, excision of the Ds element can
result in patches of cells (each progeny of a single cell)
expressing the colored phenotype. This is referred to as
variegation. If the Ac locus moves to a position adjacent to Ds,
it promotes its movement away from the C locus during the
development of the kernel and the locus reverts to normal
expression. The larger the number of Ac factors present, the
greater expression of variegation in the tissue. Larger coloured
patches are formed when Ds is transposed early in development;
smaller patches when Ds is transposed late.
MUTATION OF C LOCUS,a gene required
for synthesis of a purple pigment in the aleurone (a),
takes place when Ds moves into the locus (b). The
mutation disables the gene, the pigment is not made and
the aleurone is colorless. If Ac is present in the
genome, however, it promotes the transposition of Ds
away from the locus in some cells during kernel
development (c). The mutation reverts when the element
leaves, giving rise to cells in which the C locus
is functional. Each such cell gives rise in turn to a
pigmented sector in the aleurone.
In 1983, Barbara McClintock
won the Nobel Prize in Physiology and Medicine for her
discovery of mobile genetic elements (ie. transposons).
For an insightful look into not only what Dr.
McClintock did, but also, for how she thought, see:
Genetic deletions are the irreversible loss of loci, and
often result in loss of function for the cell or organism
Duplications are the increasing of certain genetic material,
and their effects are more varied
Changes in gross chromosomal structure can be caused by
double stranded breaks, ring chromosomes, breakage-fusion
bridge cycles, and more
Transposons are elements that can insert themselves into
different places in the genome - in McClintock's experiments,
the Ac and Dc loci are both required for transposition. In
this system, Ac produces the transposase that enables Dc to be
transposed.

 Deletions
and duplications are identified both cytologically and molecularly
Deletions
and duplications are identified both cytologically and molecularly
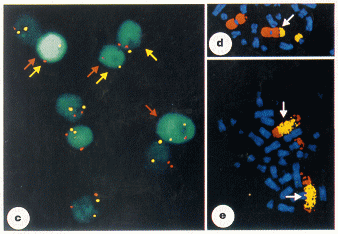
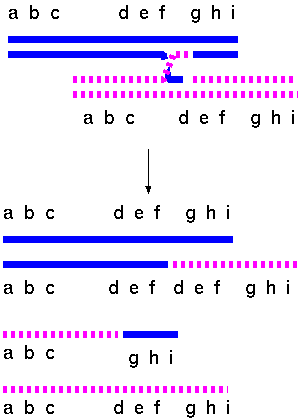
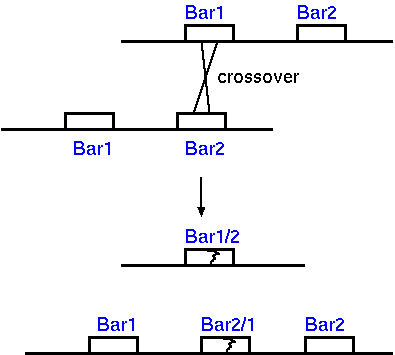
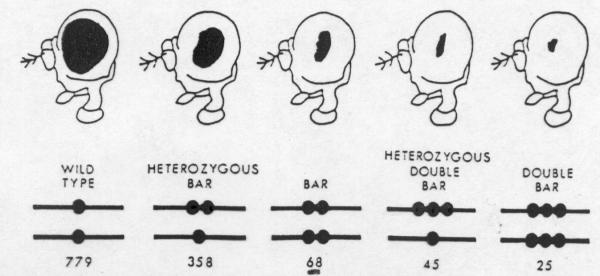 Fig. 12.6. Illustration of
the different sizes of compound eyes of the female Drosophila
melanogaster as caused by the varying numbers of facets.
The size of the eye is influenced by the position effect. (From
Kin, 1965. Redrawn from Oxford University Press, New York).
Fig. 12.6. Illustration of
the different sizes of compound eyes of the female Drosophila
melanogaster as caused by the varying numbers of facets.
The size of the eye is influenced by the position effect. (From
Kin, 1965. Redrawn from Oxford University Press, New York).