When I first began teaching this course in 1990, it was targeted at plant breeders. The course was therefore highly practical in its spin on biology. Plant breeders were interested in the fact that changes could occur in chromosome structure primarily with respect to the deleterious effects of gross structural changes on fertility. Therefore, the material was presented primarily as a set of facts to be learned, and mechanisms to be understood. The terms "evolution" and "speciation" were used sparingly, if at all.
As I began to understand these mechanisms better myself, I began to realize the profound implications that chromosomal changes had on speciation, and therefore, on the larger process of evolution. Each time I reexamined another piece of this subject matter in an evolutionary context, it suddenly clicked. It now made a great deal of sense. In my mind, a dry and convoluted area of genetics suddenly became one of the great keys to my understanding of evolution.
We've looked at transposition in the previous reading, which is the change in position of an element on the same chromosome. Translocation, by comparison, is the change in location of a genetic element, to a different chromosome.
As displayed, there is no net change in genetic information, only a change in the location of that genetic information.
There are a few different ways that translocations can occur. A couple examples are:
| Mostly, translocations show up when
chromosomes have to pair. Normally, each chromosome can pair
along its length to the homologous chromosome - but when a
translocation has occurred, pairing becomes more
complicated.
It's best to start by reviewing chromosome pairing at
meiosis. At pachytene, homologous chromosomes are
completely paired. After desynapsis during diplotene,
chromosomes remained paired at their termini. During
metaphase I, sister chromatids are still together, but
kinetochores have separated.
|
| One
way
or the other, homologous sequences will find each other at
pachytene, regardless of which chromosomes they happen to
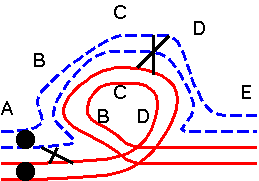
be on. When a reciprocal translocation is in the heterozygous state, the four translocated chromosomes must pair together, bending in such a way as to bring together all homologous regions, regardless of which chromosome they are on. For reciprocal translocations, a cross-shaped pairing, referred to as a quadruple, is required to bring together all homologous regions. Note: When a translocated chromosome is homozygous, no aberrant pairing occurs! |
At metaphase I, several types of configuration are possible,
depending on the segregation of the kinetechores. These
orientations are given specific designations - alternate
and adjacent - and the consequence for the
translocation heterozygote are distinct.
|
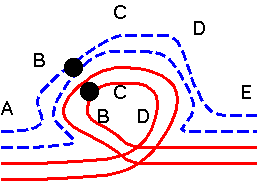
Alternate: both normal chromosomes go to one pole, both translocated chromosomes go to the other pole. Result: balanced gametes. |
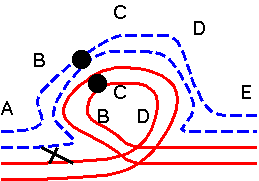
| Adjacent :
One normal and one translocated chromosome move to each
pole. Result: unbalanced gametes When sister chromatids for a translocated chromosome segregate to opposite daughter cells, both daughter cells are missing part of one chromosome and have a duplication for another part. When sister chromatids for a translocated chromosome segregate to opposite daughter cells, both daughter cells are missing part of one chromosome and have a duplication for another part. |
Adjacent-1: One normal and one translocated chromosome move to each pole (normal behavior for centromeres).
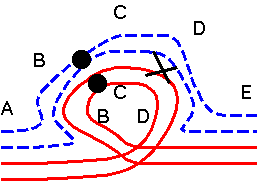
Adjacent-2: Both homologous centromeres move to the same pole (rare, abnormal behavior for centromeres).
These combinations result at least in an equal number of
chromosomes at each pole, if not equal genetic information. In co-orientation
configuration, the chromosomes are distributed in equal
numbers to opposite poles. In adjacent-1, the homologous
kinetechores move to opposite poles. However, sometimes the
chromosomes do not split equally between the poles.
|
Non-coorientation : two non-co-oriented chromosomes are stretched across the metaphase plate and not attached to the poles. Result: unbalanced gametes At anaphase, the segregation is 1 cooriented and 2 non-cooriented chromosomes migrate to one pole, and 1 cooriented chromosome to the other pole producing a 3:1 chromosome segregation which results in aneuploid progeny-trisomic and monosomic.
|
The region between the kinetechores and the translocation is called the interstitial region. If no cross-overs occur in the interstitial region, alternate chromosome migration results in balanced gametes: one half containing the normal chromosomes 1 and 2, the other half with the 1' and 2' translocated but balanced chromosomes. The resultant meiotic products depend upon how the kinetochores segregate.
| |
Alternate | Adjacent |
|---|---|---|
| Crossover between centromere and translocation breakpoint. | ||
| Crossover after translocation breakpoint. |
There is an approximate 1:1 ratio of alternate to adjacent arrangements of chromosomes in meiosis of an interchange heterozygote.
The consequences of crossing-over varies. In diploid plants, the presence of a heterozygous interchange produces approximately 50% pollen sterility. This is due to the 1:1 ratio between alternate (viable ) and adjacent (non-viable) orientations. In polyploids, interchange heterozygotes do not show 50% pollen and seed sterility although the parents may differ by two interchanges. Male and female gametes containing dup/del in polyploid are as competitive as normal gametes due to the genetic compensation possible in polyploids.
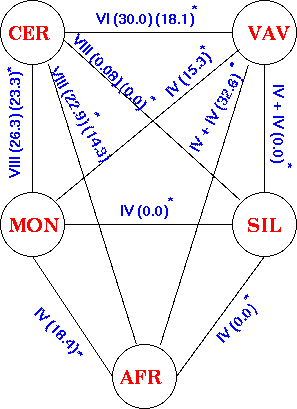
If a primitive species can be identified, chromosome pairing can be observed in crosses with related species to discover how chromosome structural changes have led to speciation. Let's use as an example the Secale rye.
The cultivated species is Secale cereale rye and four wild species: S. vavilovii, S. africanum, S. montanum, and S. silvestre. All contain 2n=14 chromosomes. Reciprocal translocations have contributed to the effective reproductive isolation of the wild and cultivated species. Translocations set limits on genetic exchange because of the resulting sterility.
Normal chromosomes should give the characteristic "D" shape appearance at meiosis. When chromosomes in a hybrid have undergone a reciprocal translocation, the translocated ends will be paired in the hybrids at the end of prophase I. The result is chains of chromosomes.
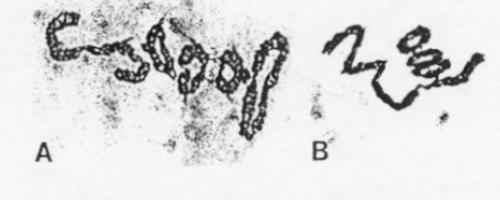
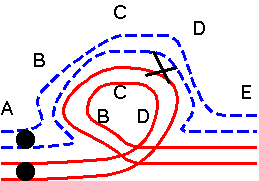
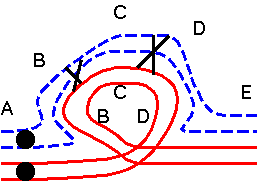
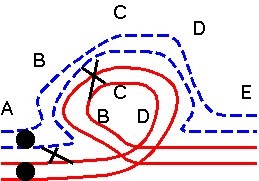
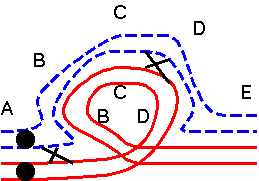
The results of hybridizing diferent Secale species are summarized at right. Because homologous arms pair at pachytene and are still attached after terminalization at diakinesis, chromosomes enter metaphase I in chains, in which homologous ends are linked. Redrawn from Figure 6.14. Cytogenetic relationships among five major Secale species. Abbreviations: AFR, africanum ; CER, cereale; MON, montanum; SIL, silvestre ; VAV, vavilovii. Crossability rate (%) in parentheses is from Khush (1962) and in parentheses with an asterisk is from Singh (1977). Roman numerals: The number of chromosomes that pair as a chain. |
|
The pairing relationships in parents and hybrids can be represented in a diagram.
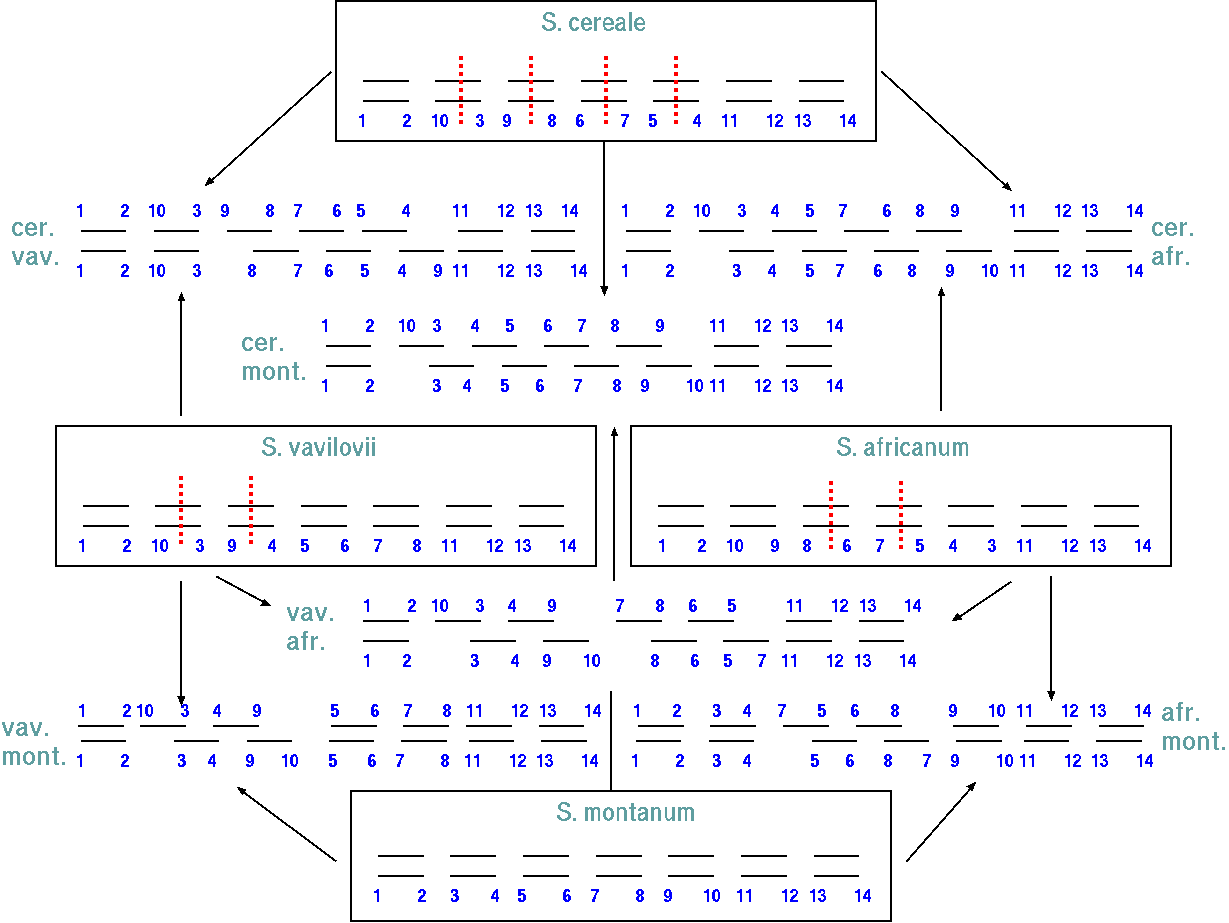
| Inversions are changes in linear sequence which result in the reverse order. Inversions are the most common type of chromosome aberration found in natural animal and plant populations, and are most common in metacentric and submetacentric chromosomes. The causes are natural and induced chromosome breakage, followed by reunion in the reverse orientation. In the formation of an inversion, chromosome segments can be separated from their mother chromosomes by breaks and then reinserted into another homologous chromosome in reverse order. No duplication is involved. Inversions also contribute to speciation. They have no effect on mitotic divisions but do affect meiosis. In a cell heterozygous for an inversion, pairing cannot occur in a linear fashion and pairing requirements can be met by the inverted section forming an inversion loop. This inversion loop causes problems during meiosis when a crossing-over event occurs. |
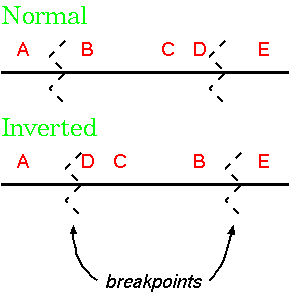 |
The two types of inversions are:
Small inversion loop on chromosome 2 in mice (arrow). Pachytene chromosomes are visualized using antibodies to scaffold protein SCP3 (magenta). Meiotic Exchange and Segregation in Female Mice Heterozygous for Paracentric Inversions Kara E. Koehler1, Elise A. Millie, Jonathan P. Cherry, Stefanie E. Schrump and Terry J. Hassold doi: 10.1534/genetics.166.3.1199 Genetics March 1, 2004 vol. 166 no. 3 1199-1214 Displayed by link to https://www.genetics.org/content/genetics/166/3/1199/F3/graphic-6.large.jpg |
Pachytene
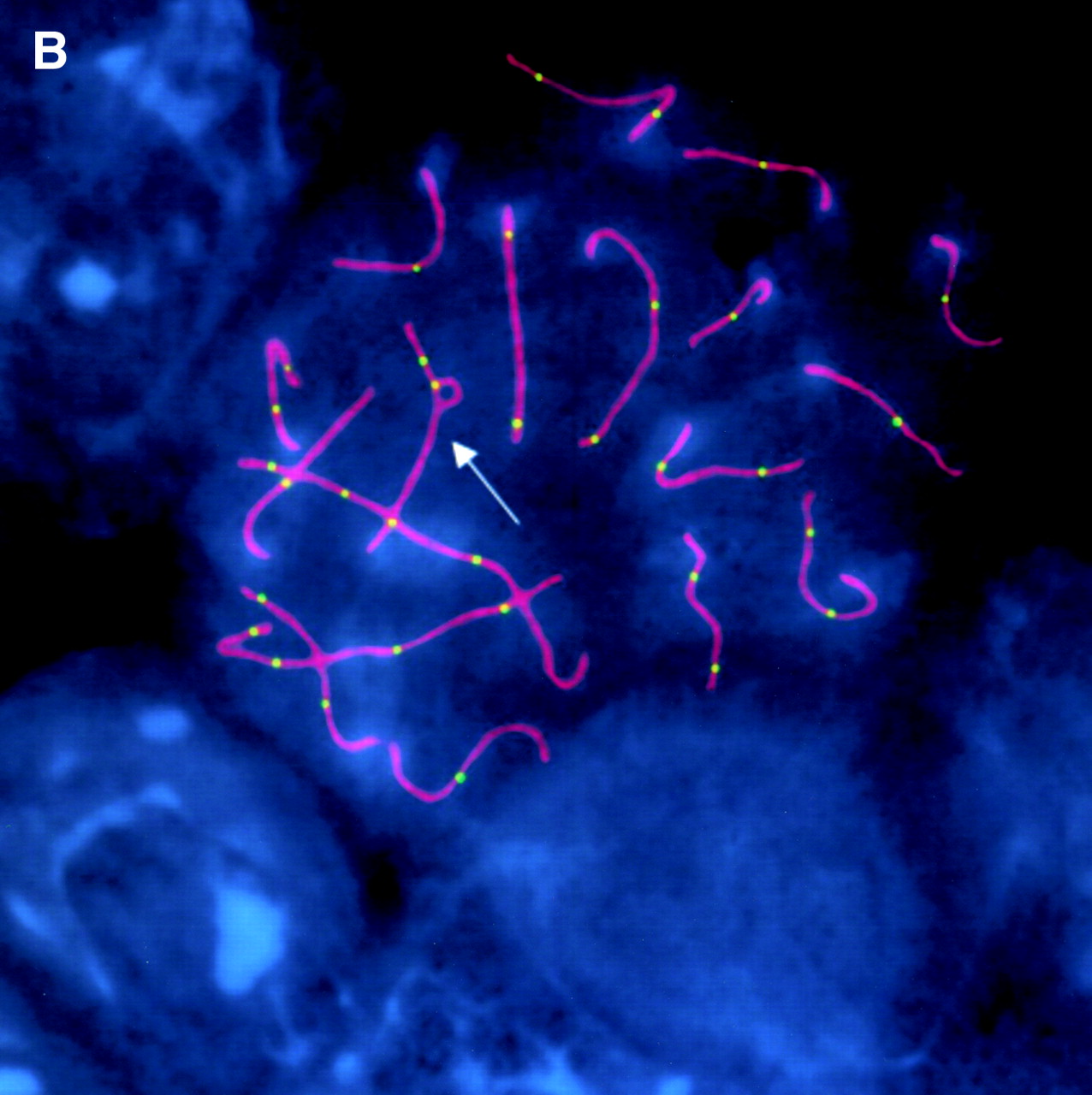 |
Both breaks occur in the same arm so the inverted region does not include the centromere. Paracentric inversions are observed more frequently than pericentric inversions.
In an inversion heterozygote, pairing of the inverted chromosome with the original chromosome requires a loop at pachytene. If crossovers occur within the inversion loop during pachytene, bridges and acentric fragments will be seen in anaphase I and II of meiosis.
Different configurations of dicentric bridges and fragments are dependent on the types of crossover within the loop.
An inversion loop in the A arm of a polytene chromosome from an Axarus species midge Arrow shows the location of the centromeric band. http://en.wikipedia.org/wiki/File:A_arm_inversion.jpg This is a file from the Wikimedia Commons. |
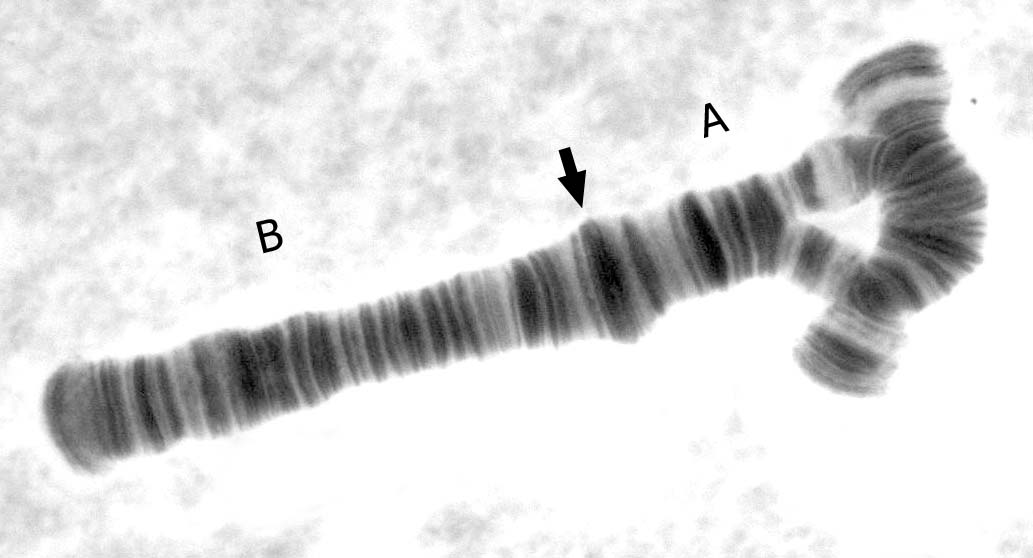 |
Note: Inversion has no effect in homozygotes. Abberrant
pairing is only seen in heterozygotes.
| Paracentric Inversions |
|
| No crossover: 2 normal and 2 inverted chromosomes |
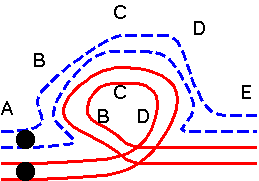 |
| Single crossover outside inversion: 1 normal and 1 inverted chromosomes, 2 recombinant chromosomes |
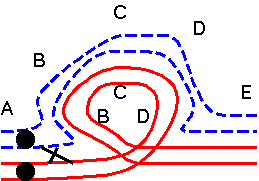 |
| Single crossover within inversion: 1 normal and 1 inverted chromosome 1 dicentric and 1 acentric fragment |
 |
Double crossovers
| 2 strand DCO - 2 strand DCO -
the same two chromatids participate in both crossover
events; both crossovers occur within the inversion loop 2 normal and 2 inverted chromosomes |
 |
| 2 strand DCO - the same two
chromatids participate in both crossover events; one
crossover is outside the inversion loop 1 normal and 1 inverted chromosome 1 dicentric and 1 acentric fragment |
 |
| 3 strand DCO - one chromatid
participates in crossovers with two different chromatids 1 normal and 1 inverted chromosome 1 dicentric and 1 acentric fragment |
 |
| 4 strand DCO - each of the
two crossovers involve two different chromatids, so that all
four chromatids are involved single crossovers 1 normal and 1 inverted chromosome 1 dicentric and 1 acentric fragment |
 |
A pericentric inversion results in morphological change in the chromosome as the centromere position and arm ratio are both altered. Depending on size of inversion, different meiotic pairing configurations may be formed in an inversion heterozygote. If the inversion is small, there will be no pairing in the inversion region. If it is large, the result is the formation of an inversion loop involving most of the chromosome. Only the region within the inversion will pair, leaving the ends unpaired. In terms of the effect on gametes, if the inversion is small there will be no further changes. If the inversion is large, crossing over can result in unbalanced recombinant chromosomes.
| Pericentric Inversions |
|
| No crossover: 2 normal and 2 inverted chromosomes |
 |
| Single crossover outside inversion: 1 normal and 1 inverted chromosomes, 2 recombinant chromosomes |
 |
| Single crossover within inversion: 1 normal and 1 inverted chromosome 2 dp/df chromosomes |
 |
Inversions don't prevent crossovers, per se. However, gametes from an inversion heterozygote will be more likely to cause unbalanced chromosome complements in zygotes, which will decrease viability of progeny. Therefore, progeny with crossover events will be rare. We observe this in an apparent reduced frequency of recombination in inversion regions.
Crossing over inside and around the inverted segment is reduced as a result of incomplete pairing eg. very short inversion segment that does not pair. A very large inverted segment forms loops, and crossing over is reduced inside and around loop due to incomplete pairing. To be precise, crossover frequency is inversely proportional to the length of the inverted segment.
In plants, heterozygous pericentric inversions cause pollen and ovule abortion because spores are produced that contain chromosomes with deletions and duplications.
The terms "evolution" and "speciation" are often misused,
despite the fact that these terms are critical to our
understanding of almost every area of biology. Evolution can be
thought of as a broad umbrella term that encompasses all
processes by which genetic changes occur at the population level
over time. Speciation is one aspect of evolution that is
both a result of evolution, and an engine for further evolution.
Evolution is a change in allele frequency within
a population over time. This is the definition favoured by
population geneticists. In the broad sense, if we define the term
"allele" to include any heritable genetic change, including
mutations in non-coding DNA, and changes in chromosome
organization of structure, including translocations, deletions,
insertions, inversions, polyploidies and aneuploidies.
Speciation is the division of a population into two subpopulations, as a result of heritable reproductive barriers. Reproductive barriers include any heritable mutation or change in chromosomal structure that would prevent mating or result in infertile offspring produced by a cross between members of the two populations.
Based on what we have discussed in this section, it should be
obvious that any mechanism, such as translocations or inversions,
that can interfere with meiotic pairing would establish a powerful
reproductive barrier, leading to speciation.
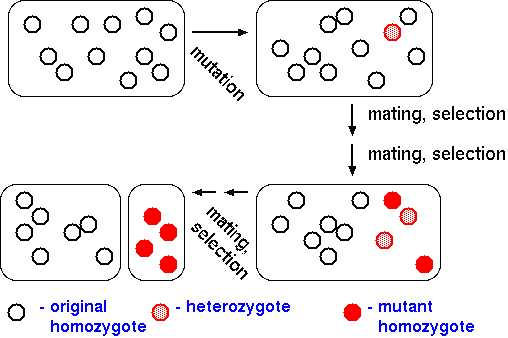
When a mutation such as a translocation occurs, it begins as a single mutational event in a single individual. When this individual mates with so-called "wild type" members of the population, some progeny may carry that mutation. In the second generation, mutant progeny may mate with each other, yielding some progeny that are homozygous for the mutation. In subsequent generations, homozygotes for the mutation will be more fertile, and thus have greater reproductive fitness than hybrid progeny. The result will be two populations between which there is minimal gene flow. As further mutations occur within each population, gene flow between the populations will cease. Once reproductive barriers arise, the two species will accumulate more mutations, thus diverging over time.
It is almost as important to explain what evolution is NOT, as it is to define the term. Evolution is best thought of as the observation that allele frequencies have changed in a population, making no judgement as to the mechanisms of the change. This is a critical nuance, because many people assume that evolution is synonymous with selection. But selection is only one of the processes that can drive evolution ie. that can cause a change in allele frequency.
In this section we will define the mechanisms that can change
allele frequencies in a population. These mechanisms are usually
categorized to include mutation, migration, selection, random
drift, non-random mating, and linkage disequilibrium. In order to
describe these mechanisms, we must first provide a context in
which to describe these changes: the Hardy-Weinberg Equilibrium.
There is an interesting parallel between Newton's First Law of
Motion and the Hardy-Weinberg Law:
| Newton's First Law of Motion Objects in motion remain in motion, and objects at rest remain at rest, unless acted upon by external forces |
Hardy-Weinberg law (HW) Allele frequencies in a population will stay the same from one generation to the next, unless other mechanisms, such as mutation, migration, selection, random drift, non-random mating or linkage disequilibrium act to change them. |
One might be inclined to dismiss Hardy-Weinberg as being
unrealistic and therefore irrelevant, because one or more of these
mechanisms will always be acting in any wild population. However,
the point of Hardy-Weinberg is to provide the theoretical
framework that allows us to measure changes in allele frequency,
as a deviation from the ideal behavior expected under
HW. That is to say, HW predicts that if the frequencies of alleles
A and a in generation 1 are p1 and q1, then the respective
frequencies in generation 2 should be p2 = p1 and q2 = q1 . If we
observe in generation 2 that p2 ¬= p1 and q2 ¬= q1, then by
definition, evolution has taken place. Once again, this makes no
judgement as to which mechanisms contributed to the change in
allele frequencies. Evolution means simply that they have changed.
The Components of Evolution
The mechanisms underlying evolution can be broken down into a number of categories, each of which can act independently and simultaneously to influence the transmission of alleles from one generation to the next.
| mutation A mutation is any heritable change in the genome of an organism. This would include point mutations such as transitions and transversions, insertions, deletions, and various sorts of rearrangements such as inversions, duplications or transpositions. Any change in structure or number of a chromosome, such as a translocation or polyploidization, counts as a mutation. |
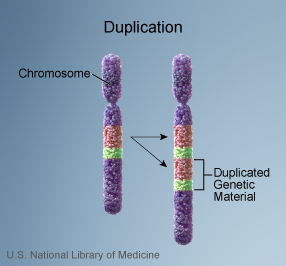 from http://ghr.nlm.nih.gov/handbook/illustrations/chromosomalduplication |
| migration
(gene flow) Individuals or gametes can flow between populations. In an extreme case, gene flow can introduce a new allele into a population that previously was not found in the population. Of course, gene flow can only occur when progeny can produce fertile offspring. For example, pollen from polyploid plant fertilize a diploid plant, the hybrid progeny will probably be sterile. |
 from http://www.slideshare.net/klemmistry101/hardy-weinberg-populations |
| selection Selection occurs when progeny exhibiting different phenotypes have different survival rates. Alleles that confer a selective advantage would be said to have a greater fitness than those that confer a selective disadvantage. Some alleles confer neither an advantage nor a disadvantage, and are said to be selectively neutral. Generally, chromosomal abnormalities, unless in the homozygous state, will result in progeny that do not have balanced numbers of genes at each locus, most likely resulting in a decrease in fitness. |
 |
| random
drift (sampling error) While it is true that HW predicts that allele frequencies will stay the same from one generation to the next, in practice, they are never precisely the same. For example, if the initial population had a perfect 50:50 mix of two alleles, there are a lot more ways to get a slightly different ratio in the next generation (51:49, 49:51, 52:48 etc.) than there are to get exactly 50:50. Once a difference exists, it can be magnified over time. Ramdom drift is most important when population bottlenecks occur, such as a limited migration of a small number of individuals to a new habitat. For example, if an initial habitat has both brown and gray mice, and small group of predominantly brown mice move to a new habitat, the brown mice will probably be the only ones found after several generations of random mating. This is not due to any selective advantage of brown over gray. Rather it is due to the luck of the draw. |
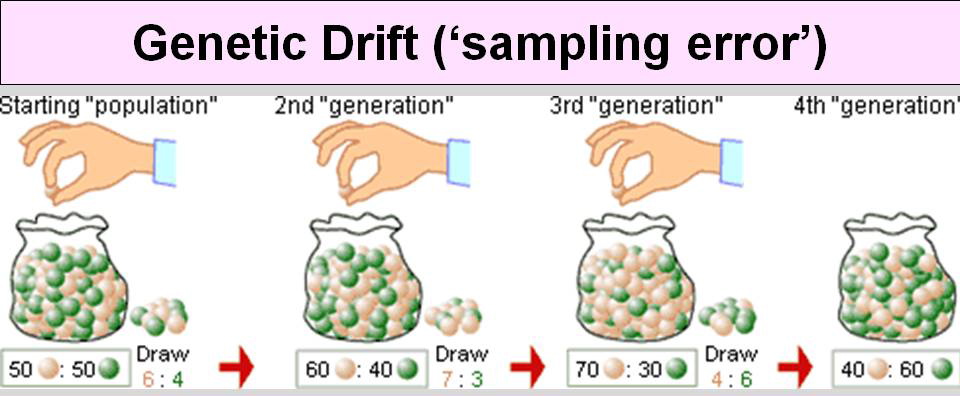 from http://uvmgg.wikia.com/wiki/Genetic_drift |
| non-random
mating Anything that biases the choice of mates in sexual reproduction can have a strong effect on gene flow between two subpopulations. Given enough time, this can lead to reproductive barriers that can result in speciation. |
 |
| linkage
disequilibrium (genetic hitchiking) When an allele confers a strong selective advantage, alleles nearby that are tightly-linked to the first gene will also increase in frequency, even though they themselves confer no selective advantage. This is because in a few generations of intense selection, there is not enough time for significant amounts of crossing over to occur between the selected gene and loci nearby. |
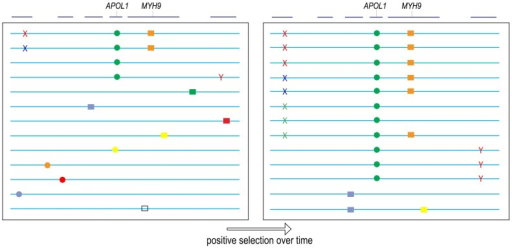 from http://openi.nlm.nih.gov/detailedresult.php?img=3530541_pone.0051546.g004&req=4 |
| To return to our physics analogy, the
components of evolution described above can be likened to
the components of a vector. In the example at right, the
apparent ground speed that we measure for an airplane is
the resultant vector whose components are the airspeed of
the plane and the velocity of the wind that alters its
path. |
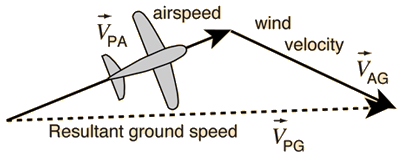 from http://hyperphysics.phy-astr.gsu.edu/hbase/airpw.html |
{kind=link}