While our task of building
a chromosome has been of enormous scope, the main thing we've done
so far is simply to describe how DNA is packaged in chromosomes,
and some of the functional ramifications of that packaging. We
still need to identify some important functional features of
chromosomes that make it possible for them to replicate and
segregate. We will describe a minimal set of features required for
normal replication and segregation. These are centromeres, origins
of replication, and telomeres.
Chromosomes need origins of replication for DNA synthesis to
occur
An origin of replication (ori) is a site at
which DNA replication begins. Origins of
replication are often AT rich sequences, which allows them to be
more easily pulled apart than GC rich sequences. Without an origin
of replication, DNA cannot be replicated and will not be passed on
to the next generation.
Eukaryotic chromosomes contain numerous
replication origins, allowing for DNA synthesis to proceed
in parallel at many sites simultaneously. Each pair of
replication forks is referred to as a "replicon". Replicons
can be visualized as bubbles of replicated DNA that expand
in both directions. Eventually, all replicons merge into a
single large bubble, until replication terminates at the
telomeres.
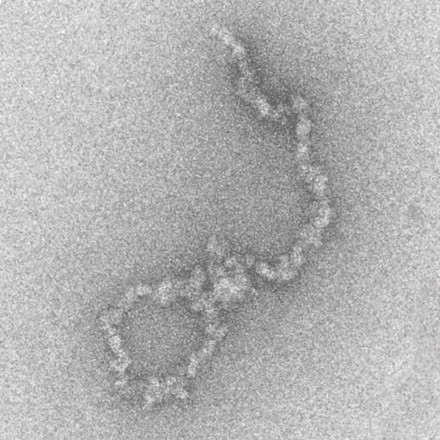
The electron micrograph at right shows several replicons,
each originating from a different origin (arrows).
Each replicon has two replication forks, moving in opposite
directions. Ultimately, replication forks meet, until
replication of each template strand is complete.
A series of experiments from Jack
Szostak's lab helped to define the critical components of
chromosomes. These experiments all had the same steps:
1. Transform yeast, deficient in Leucine
production (Leu-) with an artificial construct
2. Isolate a colony on complete media (permissive conditions)
3. Grow in complete media for several generations
4. Transfer to minimal media without Leucine
5. Identify clones that grow without Leucine
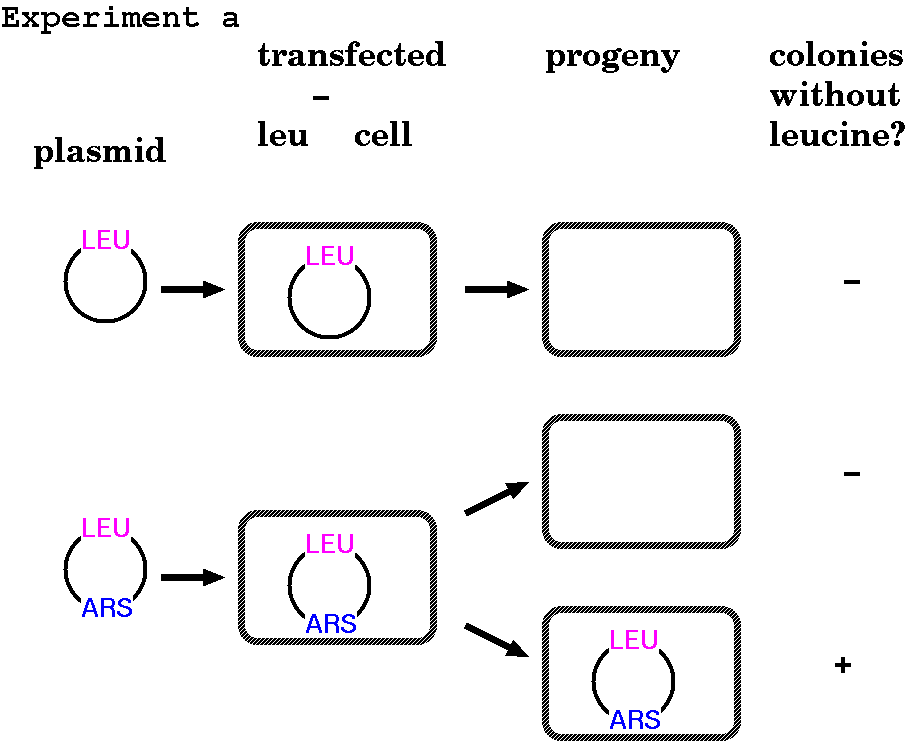
FUNCTIONAL
CHROMOSOMAL ELEMENTS: EXPERIMENT A - Origins of
Replication
If you transfect into yeast
a plasmid containing a selectable marker, in this case a
LEU gene (Leucine biosynthesis) into Leu-
yeast, the cells will not grow, even though you have
given them the correct gene. But if you randomly
clone yeast sequences into this same plasmid and
select on minimal media, you can recover a few clones
that are able to grow (ie. synthesize leucine).
The inserts contained in these surviving plasmids are
origins of replication, referred to in yeast as autonomously
replicating sequences (ARS).
ARS have been identified in a
variety of other species (eg. humans, Drosophila, maize,
tobacco, even bacteria) by virtue of their ability to
replicate in yeast.
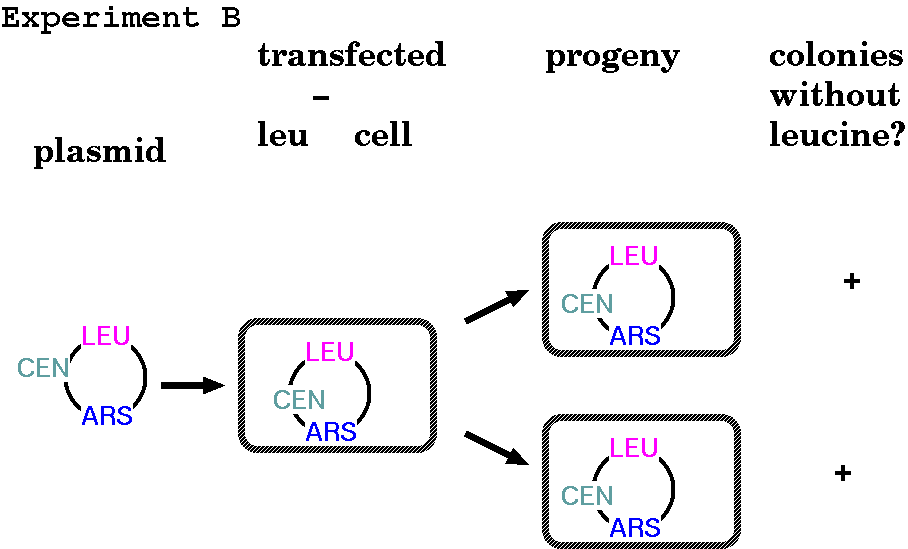
Centromeres ensure equal division of genetic material between
daughter cells
FUNCTIONAL
CHROMOSOMAL ELEMENTS: EXPERIMENT B - Centromeres
You might have noticed in the previous diagram that in the
case of the ARS plasmid transformants, not all the daughter
cells received the construct. Furthermore, in the absence of
selection, even those cell lines may lose the construct.
When yeast genomic DNA is randomly cloned into plasmids
containing both a LEU gene and an ARS, some are stably
inherited in the absence of selection. These plasmids
happened to receive a fragment of DNA containing a centromere.
Centromeres are sequences of DNA to which the
kinetochore proteins attach, and therefore to which spindle
fibres attach during mitosis. Without a centromere, the
construct has no guarantee of being passed on to both
daughter cells during cell division. They tend to include
repetitive sequences (satellite DNA), and also to not have
many nucleosomes. Presumably, that is to make spindle fibre
attachment during mitosis easier.
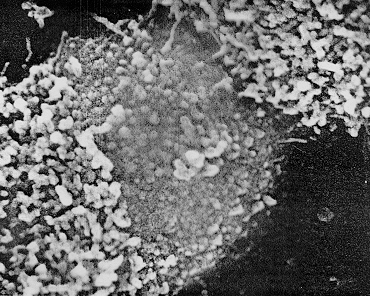
Detail of a positive staining centromere
region of a C-banded human chromosome. (C banding
preferentially stains constitutive heterochromatin). The
fibrous organization of this region is still apparent but
is either covered by, or embedded in, an amorphous matrix.
Magnification x 64,000. Chromosomes and Chromatin, Vol. II
(1988) Ed. K.W.Adolph. CRC Press. Fig. 14 pg. 67
Telomeres are needed for completion of replication, and
protection, of the ends of chromosomes
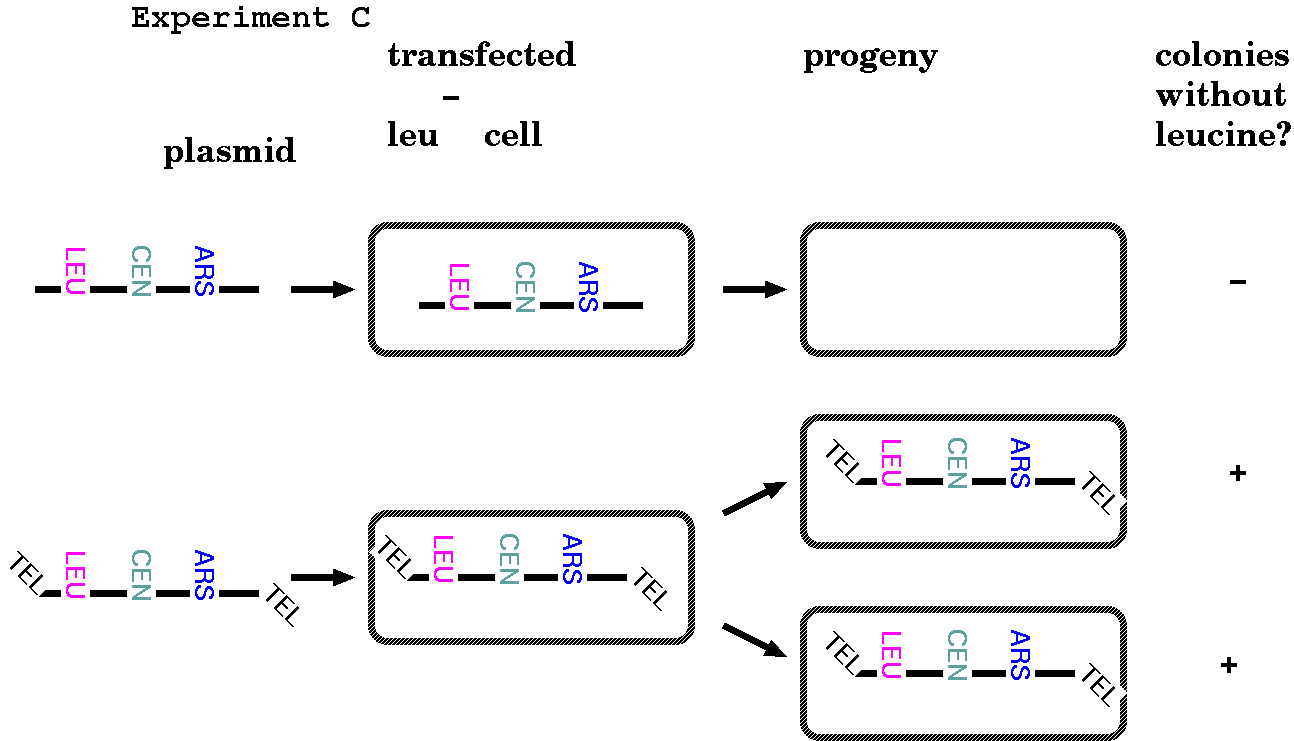
FUNCTIONAL
CHROMOSOMAL ELEMENTS: EXPERIMENT C - Telomeres
In the first two experiments, the researchers created what
are now known as Yeast Artificial Chromosomes (YACs). Up to
now, we've been getting away with creating artificial
chromosomes by making them circular. In yeast, they function
as completely independent chromosomes. But eukaryotic
chromosomes are linear, and Experiment C below illustrates
that linear chromosomes can't normally replicate without
telomeres at each end.
When Tetrahymena telomeric
sequences were added to the ends of linearized yeast
artificial chromosomes, the YACs were able to replicate
and segregate stably in yeast.
Why is that? What function do telomeres serve that make them so
necessary for linear chromosomes?
The basic problem with linear chromosomes are
the ends - specifically, the 5' ends of a new strand. DNA
polymerases can synthesize from 5' to 3' only. While the
leading strand can read right to the end of the template,
the lagging strand cannot. What ends up happening is this:
Parent strands are in dark blue.
As you can see, the 5' ends of the daughter strands are short.
Now, for one replication, this might not matter so much. The
daughter cells would lose some DNA off the chromosome ends, but
not too much. Probably, function would not be affected. But what
happens in the next division, and the next? More and more DNA is
lost and the chance of losing part of an important gene is much
more likely. Put this way, a species with linear
chromosomes and without telomeres is unlikely to survive for more
than a few generations.
Prokaryotes avoid the problem of replicating the ends of linear
molecules by instead having circular chromosomes
Circular chromosomes satisfy the requirement that there
must be an upstream DNA polymerase complex to remove RNA primers
and fill in gaps.
Telomeres are specialized sequences that facilitate the
replication of the ends of linear chromosomes and protect them
from nuclease digestion.
Linear molecules will always have an unfilled gap upstream from
the origin of replication closest to each end of the chromosome.
Since this unfilled gap will always be a 3' protruding end, there
is no way for DNA polymerase to "fill it in". Given that
eukaryotes evolved so long ago, there must be a mechanism for
dealing with this problem.
That solution is the telomere. Telomeric
sequences are composed of repeated motifs and extend the linear
eukaryotic chromosome. Think of telomeric sequences like a buffer
for loss of sequence. As long as you add roughly as many
nucleotides to the ends of chromosomes as they lose during each
round of DNA replication, there is no net loss of DNA at the ends.
Telomeric sequence data was first obtained in Tetrahymenathermophilus, a cilliated protozoan. Tetrahymena
has two separate nuclei, a germinal micronucleus, which carries a
diploid complement of chromosomes, and a somatic macronucleus, in
which particular chromosomes or parts of chromosomes are present
in very high copy numbers. The rDNA genes are present on separate
chromosomes with about 104 copies. Because the rDNA
genes are located near the ends of the chromosomes, it was
therefore possible to directly sequence the termini of chromosomal
DNA using rDNA-specific primers. These studies revealed that
individual rDNA termini carry a variable number (~120-420bps) of C4A2/T2G4
repeats.
Other examples: Oxytricha C4A4 Saccharomyces C2-3A(CA)1-3 Dictyostelium C1-8T H. sapiens CCCTAA
These telomeric sequences are oriented
so that the C-rich strand always runs 5'-->3'
from the end towards the interior of the chromosome. In some
cases, there are nicks, or gaps in the C-rich strand.
Example: Oxytricha (ciliated
protozoan):
end of chromosome to centromere --> 5' C4A4C4A4C4----------------------..... 3' OH-G4T4G4T4G4T4G4T4G4----------------------.....
Telomeres actually have two functions:
protection of linear ends from degradation by nucleases
acting as a template for elongating the ends of the
chromosomes.
Flourescent-labeled
DNA fragments containing telomeric sequences were
hybridized to metaphase chromosomes. Telomeric probe is
visualized in white. Chromosomal DNA is counterstained
with DAPI (blue).
Telomeres prevent degradation, repair
and recombination of chromosome ends
The overhang ends are still a problem, though. Typically, the end
of a linear DNA molecule would be recognized as a damaged piece of
DNA and "repaired", resulting in loss of sequence from the ends of
chromosomes, or fusion with other double-stranded DNAs. Eukaryotic
cells have mechanisms for protecting chromosome ends from repair
enzymes, which vary among eukaryotes. For example, telomeres of
ciliates and fungi are protected by telomere binding proteins
which effectively hide the telomeres from repair machinery.
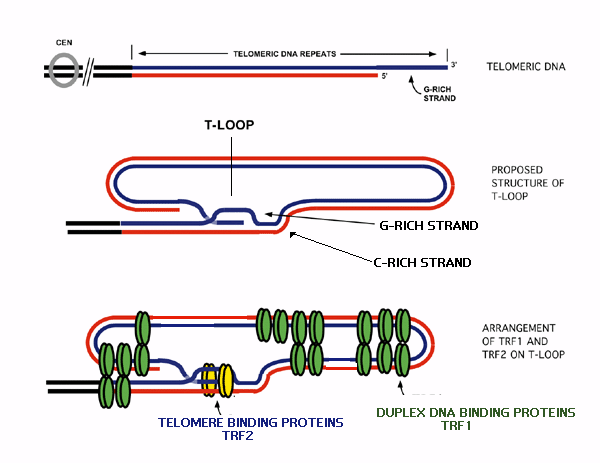
Mammals have a more elaborate D-loop
structure, in which the double-stranded telomere DNA opens
up to form a single-stranded D-loop (or T-Loop). The 3'
protruding end can loop back to form a T-loop. The end of
the T-loop can base pair with internal repeat units by
non-Watson-Crick base pairing. The D-loop is maintained
through additional proteins that bind both the T-loop and
the D-loop seen in the figure.
Telomerases carry a small RNA molecule which
acts as a template for elongation of telomeric repeats.
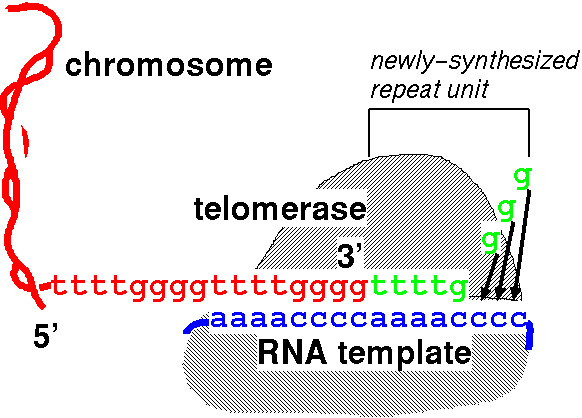
Functionally,
the telomere is a 3' protruding end which can not be
elongated by DNA polymerase. Telomere terminal transferase
(telomerase) allows the telomere to function as a 3'
recessed end in the following way: The telomerase enzyme
carries an RNA molecule (blue) whose sequence is the
complement of the telomeric repeat. When the telomere (red) base pairs with this RNA,
the 5' end of the RNA extends beyond the end of the
telomere. This allows the RNA to act as template for
the addition of nucleotides (green)to
the telomeric 3' end, which acts as a primer. In the
example, an RNA template with the sequence 5'(ccccaaaa)n3'
codes for the actual telomeric repeat 5'(ttttgggg)n3'.
So, by extending the 3' ends
of linear chromosomes with each round of DNA synthesis,
telomerase provides a longer template for the lagging
strand, which offsets the inevitable loss of DNA from
the ends.
What do you think came first - telomeres or linear
chromosomes? Why?
Telomerase activity varies with cell type and developmental
stage
Is telomerase active in somatic cells? As it turns out, the
answer depends on species is being observed, and stage of
development. Below is an incomplete list of organisms along with
telomerase activity in their somatic cells. Clearly there is much
variation between eukaryotes when it comes to somatic telomerase
activity.
Is telomerase active in
somatic cells?
Unicellular eukaryotes
- Telomerase is required in each cell division to maintain
telomere length
Humans1 - Telomerase activity
is usually only seen in stem cells or germline cells, and
telomerase activity is usually not found in somatic cells.
It is hypothesized that because of the long human
lifespan, somatic suppression of telomerase activity
occurs as a check on cell proliferation, which could
otherwise result in cancer. This is not a perfect control,
because ultimately as telomeres are lost, oncogenes near
the telomeres begin to be lost as well, resulting in
cancer.
Mice1 - Telomerase activity
is often found in somatic cells in mice. This observation
makes sense in contrast to the lack of telomerase activity
in human somatic cells, because mice have very short life
spans, and therefore would have less need for suppressing
telomerase activity as a way of suppressing cancer.
Drosophila3
- doesn't use traditional short telomeric repeats
elongated by telomerase. Instead, two retrotransposons,
HeT-A and TART transpose specifically to chromosome ends,
elongating the array of transposon repeats at the
telomeres. (Weird or what?)
Tobacco2 - High levels
of telomerase activity was seen in actively dividing cells
(roots and flowers), with low levels of activity in stems,
and no detectable activity in mature leaves. This is
consistent with the hypothesis that telomerase activity is
needed in rapidly dividing tissues.
1 Wong JMY and
Collins K (2003) Telomere maintanence and disease.
The Lancet
362:983-988.
2 Yang SW, Jin ES, Chung IK, Kim WT (2001)
Expression of telomerase activity is closely correlated
with the capacity for cell division in tobacco plants. J. Plant Biol.
44:168.
3
Danilevskaya ON, Traverse KL, Hogan NC, DeBaryshe GP and
Pardue ML (1999) The two Drosophila telomeric transposable
elements have very different patterns of transcription. Mol. Cell. Biol.
19:873-881.
Some prokaryotes have linear chromosomes with telomere-like
structures
Hinnebusch J, Tilly K (1993)
Linear plasmids and chromosomes in bacteria. Mol. Microbiol.
10:917-922
Volff J-N, Altenbuchner J
(2000) A new beginning with new ends: linearisation of
circular chromosomes during bacterial evolution. FEMS Microbiology
Letters 186:143-150.
Ravin NV, Kuprianov VV,
Gilcrease EB, Casjens SR (2003) Bidirectional replication from
an internal ori site of the linear N15 plasmid prophage. Nucl. Acids Res.
31. DOI: 10.1093/nar/gkg856.
Surprising, right? Why are we talking about prokaryotes - don't
they have circular chromosomes and therefore have no need for
telomeres? In biology, every rule has exceptions. There are a
number of cases in which prokaryotes have either linear
chromosomes, or linear plasmids. One well-known
example is bacteriophage Lambda, which is linear in its
encapsidated form, but circularizes, by annealing of its "sticky
ends", prior to replication as a circular molecule. However, there
are now many examples of linear chromosomes and plasmids,
including the spirochete Borrelia, the actinomycete Streptomyces,
and the plant pathogen Agrobacterium.
While linear chromosomes or plasmids are rare amongst
prokaryotes, their occurence in widely diverse species suggests
that linearity has arisen independently numerous times over the
course of microbial evolution. Two common schemes have been seen
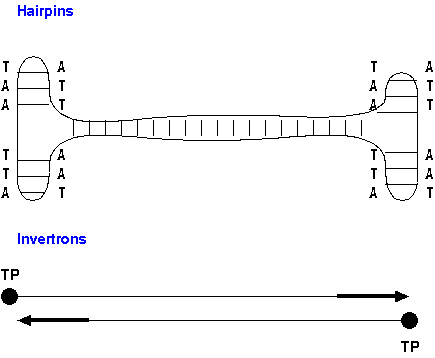
for linear chromosome replication: hairpins and invertrons.
Hairpins (eg. Borellia) - Normally linear
chromosomes contain inverted repeats at each end, which are
capable of forming hairpin loop by intra-strand base
pairing. When the leading strand from an internal
replication origin arrives at the hairpin, the hairpin
allows the template strand to be replicated in much the same
way as a circular plasmid, such that the leading strand is
redirected to "follow behind" the lagging strand. Thus,
there is always a polymerase complex upstream from each
lagging strand.
Invertrons (eg. Streptomyces) - Linear chromosomes
contain inverted repeat units at both ends. Inverted repeats
are bound by terminal proteins (TP) which bind to the 5' end
of the repeats. The terminal proteins themselves act as
primers, binding DNA polymerase. The first nucleotide to be
added to the template is covalently bound to the TP, and the
chain is elongated by further addition of nucleotides to the
3' end of that nucleotide.
Compare and contrast the methods used to deal with
linear chromosomes by prokaryotes and by eukaryotes. Do you find
that there is an underlying reason for the differences?
Summary
Chromosomes cannot replicate without an origin of
replication
Centromeres ensure that the construct or chromosome is
heritable even without selection, by providing stable
attachment sites for spindle fibres for segregation at mitosis
and meiosis
Linear chromosome require telomeres to replicate and protect
their ends by multiple processes, including D-loops, hairpins,
and telomerase action

 Parent strands are in dark blue.
Parent strands are in dark blue.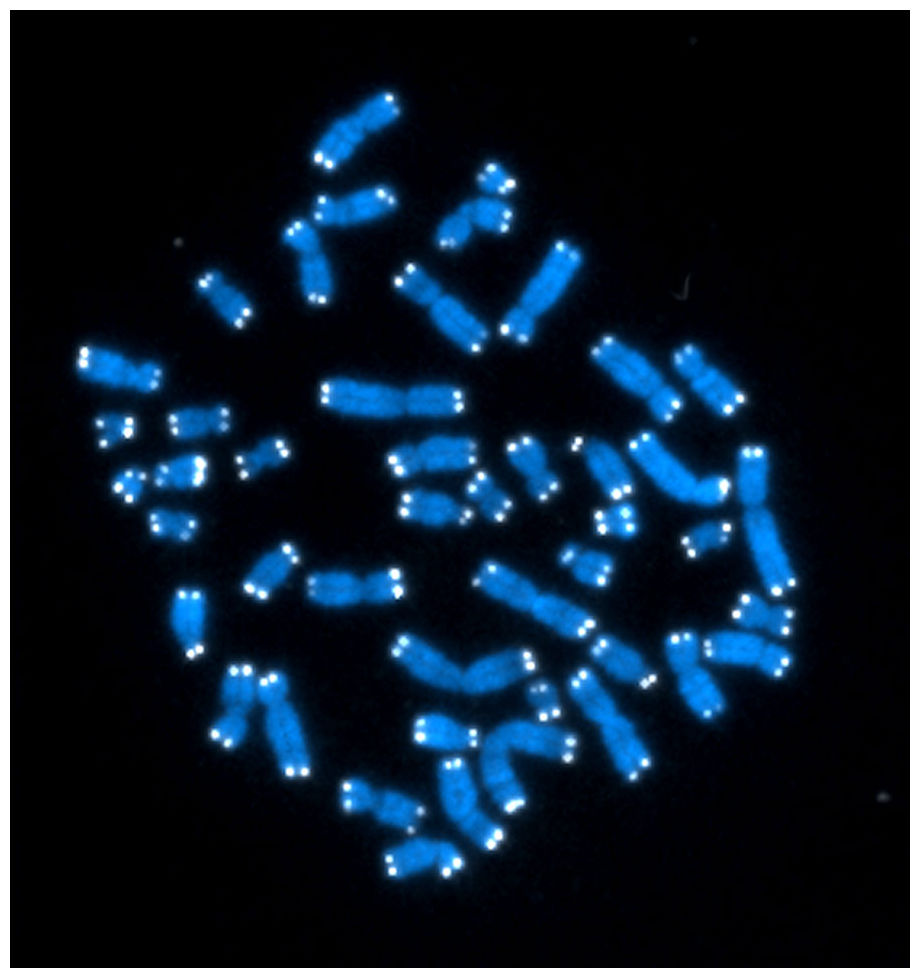
{kind=link}
{kind=link}