|
Laboratory
of RNA Biology
|
| Search | Maps | UM_IntraNet | |
|
|
|
|
|
|
|
| RESEARCH: Control of alternative pre-mRNA splicing in biology and diseases Alternative
splicing is a common way
of gene regulation that allows the generation
of multiple mRNA and often protein isoforms
from a single gene. Almost all human gene
transcripts are alternatively spliced and some
are known to generate extremely diverse
protein isoforms. This greatly contributes to
the proteomic
complexity, particularly in
neurons and endocrine cells. In
experimental animals, genetic deficiencies in
alternative splicing factors result in
developmental defects or embryonic-lethal
phenotypes. In humans, aberrant
splicing can be resulted from 15-30% of genetic
mutations that cause diseases. Thus, it is
important to understand alternative splicing
and its regulation in biology and
diseases in the omics era.
For example, in electrically excitable
cells such as neurons, endocrine and muscle
cells, ion channels allow ions in/out of the
cell membranes to generate electrical firing
patterns that are important for cell functions
including memory, hormone secretion and muscle
contraction. These processes are believed to
be critical for higher order phenotype such as
learning, behavior, metabolism and heart
beating. How these processes are finely tuned
during development and in adult life is still
a mystery to researchers. Alternative splicing
provides a unique way to diversify proteins
and may play a critical role here. A number of
studies have indicated that alternative
splicing is involved in adaptive or addictive
changes in neurons by neuronal activity
or alcohol stimulation.
Interestingly, alternative splicing of
some ion channel genes is regulated by
membrane depolarization, the first part of an
action potential, implying a gene expression
change related to the electrophysiological
memory observed in
neurons, or hormone production in response to
experience in life (e.g. exercise
or stress).
However, the molecular basis of the splicing
regulation, particularly after recurrent
stimulation, remains largely unclear.
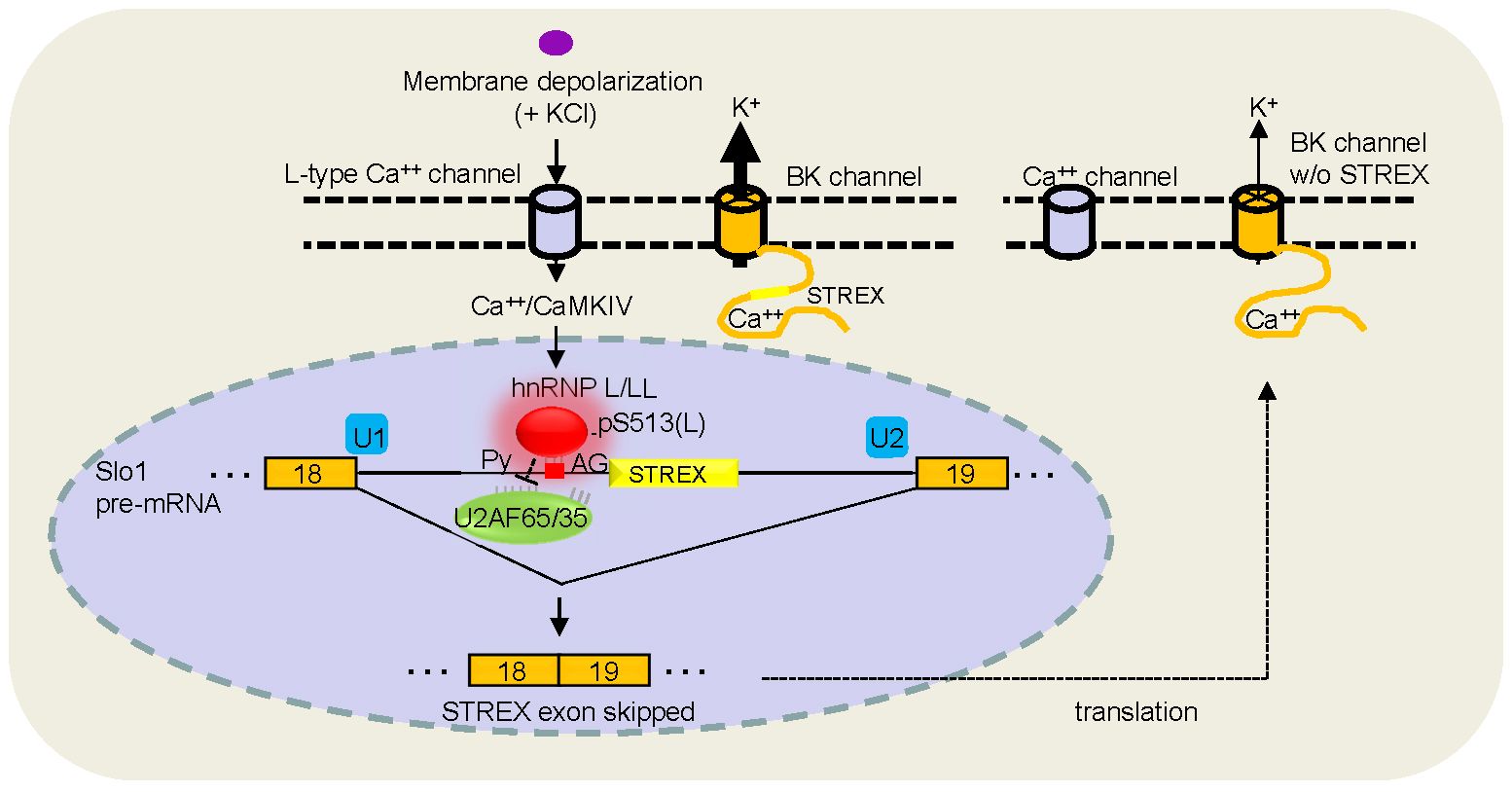
We have used the STREX (stress
axis-regulated
exon) variant of the Slo1 BK potassium
channel
gene as a model to study how cell signals
regulate the choice of alternative splice
sites in pre-mRNA transcripts. Inclusion of
the STREX exon enhances the calcium
sensitivity of BK channels and likely
modulates cellular electrical properties
related to hearing frequency tuning or
adaptive changes in learning and memory or
tolerance to alcohol. Its regulation by stress
hormones and the calcium/calmodulin-dependent
protein kinase IV (CaMK IV) makes it an
interesting target for dissecting the
components regulating alternative splicing as
well as understanding the impact of splicing
regulation on neuronal electrical properties.
A first step toward this goal was made by
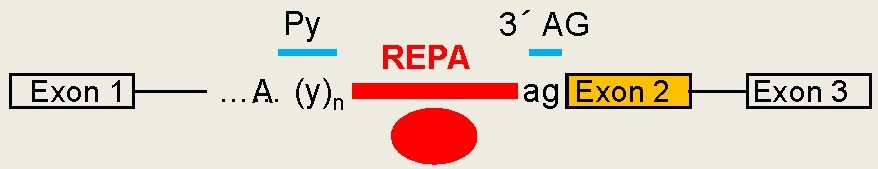
coupling CaMK IV with a pre-mRNA element (CaRRE1,
Fig. 1) sufficient to confer CaMK IV response
to an otherwise non-responsive exon. We have
recently identified the splicing factors hnRNP
L and L-Like (LL) as
essential components of the CaMKIV-regulated
splicing of STREX by inhibiting U2AF65 binding
to the upstream 3' splice site. Particularly
for hnRNP L, its Ser513 is
phosphorylated
and essential for the regulation.(Fig. 2)
Other factors including PTB and
hnRNP K are involved in
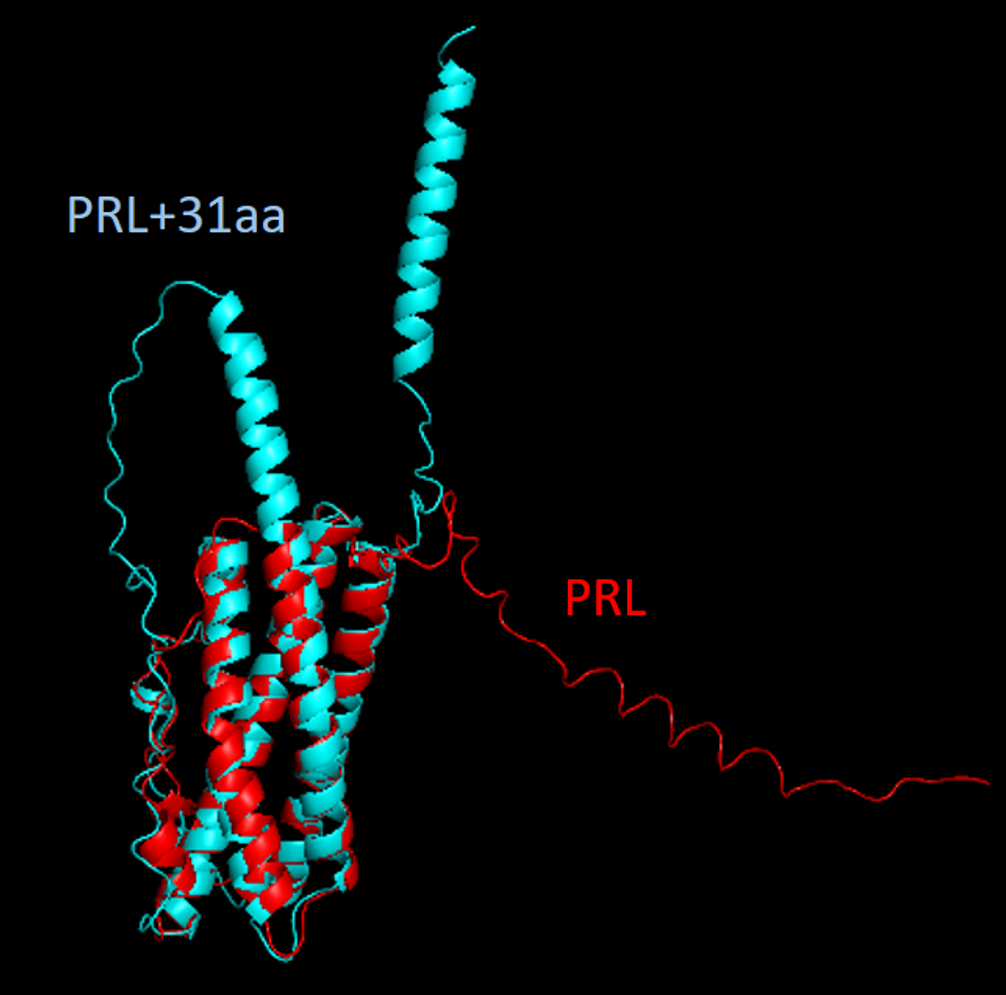
the regulation as well. In particular, hnRNP L and LL are required for the differential regulation and protection of the hormone gene expression programs for producing prolactin and growth hormones (Fig. 3). Further detailed characterization of this molecular process and its role in homeostatic or adaptive splicing (Fig 4), hormone production and stress response is ongoing .
In recent years, reports by several
labs have also demonstrated that alternative
splicing of the BK
channel and AMPA
receptor genes after chronic
inactivities or of the neurexin
gene upon depolarization/activities of
neurons plays a critical role in the homeostasis
of cellular electrical properties or synaptic
formation. Moreover, the regulation is
mediated by the Ca++/calmodulin-dependent
protein kinase IV (CaMKIV) and its downstream
splicing factors Sam68
or Nova-2,
depending on the target exons. Together, these
observations support a critical role of
depolarization-regulated splicing in hormone
production, neuronal homeostasis or
development.
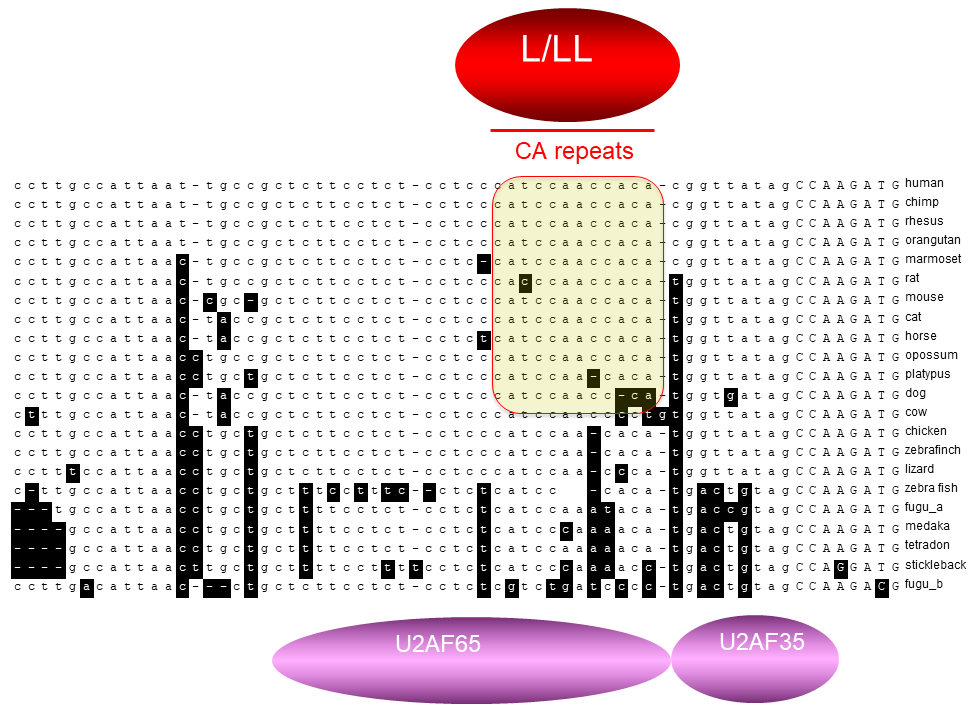
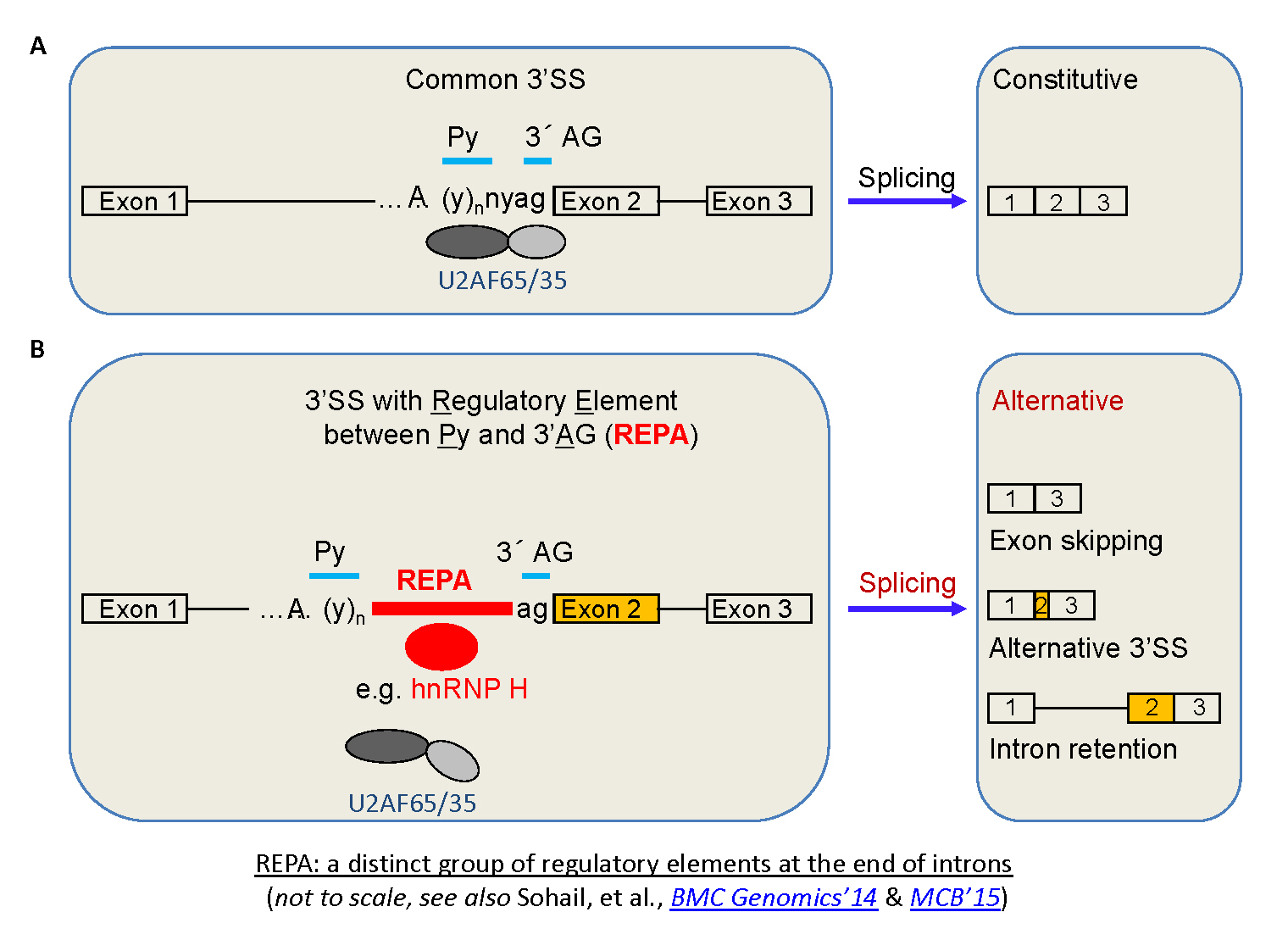
CaRRE1-like RNA
elements,
even G
tracts, have been found to act
similarly as CaRRE1 in hundreds of human
genes. Together we call these groups of RNA
sequences REPA (regulatory elements
between the polypyrimidine tract and 3' AG (Figs. 5 & 6). Most
of the REPA G
tracts (REPAG) appear to
have emerged in the ancestors of mammals,
likely contributing to the higher abundance of
alternative splicing and proteomic diversity.
We have demonstrated with the REPAG of PRMT5 exon 3
that it contributes to the evolutionary
emergence of a novel splice variant with an
opposite effect on cell cycle. The REPAGs
are widespread and highly enriched in
metazoa and plants, with the highest abundance
in mammals. They are also enriched in the aberrant 3' splice sites
of cancer patients mutated of the 3'
splicing factors SF3B1 or U2AF35. In the CBS gene, mutation of which causes
the human genetic disease
homocystinuria, a REPAG prevents its aberrant
splicing.
Other exons studied include those
involved in neuronal function,
human genetic disease or cell growth/apoptosis,
or splicing regulations involving other
signaling pathways including protein acetylation
and methylation.
Analyses of these signal-responsive RNA
elements indicate that they are mostly
mammalian-specific, likely contributing to the
more delicate and dynamic control of
alternative splicing and higher
proteomic complexity.
We hope these studies will
provide molecular details of
splicing changes in cell
physiology, as well as knowledge for cancer
diagnosis/therapy or the correction of aberrant
splicing that causes human genetic
diseases.
|
|
| PUBLICATIONS:
1.
Chunyu
Liu, Roby Joehanes, Jiantao Ma, Jiuyong Xie,
Jian Yang, Mengyao Wang, TianXiao Huan, Shih-Jen
Hwang, Jia Wen, Quan Sun, Cumhur Demirkale,
Nancy Heard-Costa, Peter Orchar, April P.
Carson, Laura M. Raffield, Alexander P. Reiner,
Yun Li, George O'Connor, Joanne M Murabito,
Peter Munson, Daniel Levy: Integrating Whole
Genome and Transcriptome Sequencing to
Characterize the Genetic Architecture of Isoform
Variation and its Implications for Health and
Disease, Nature Communications,
2025, in press, doi: https://doi.org/10.1101/2024.12.04.24318434
2.
Joyce
W, He K, Zhang M, Ogunsola S, Wu X, Joseph KT,
Bogomolny D, Yu W, Springer MS, Xie J, Signore
AV, Campbell KL: Genetic excision of the
regulatory cardiac troponin I extension in high
heart rate mammal clades. Science,
(2024); 385(6716) 1466-1471, https://www.science.org/doi/10.1126/science.adi8146. ·
Related
Perspective/Commentary: Can evolution-based
studies inform modern medicine? https://www.science.org/doi/10.1126/science.ads2585
3.
Liu
L, Nguyen H, Das U, Ogunsola S, Yu J, Lei L,
Kung M, Pejhan S, Rastegar M, Xie J: Epigenetic
Control of Adaptive or Homeostatic Splicing
During Interval-Training Activities. Nucleic
Acids Res., (2024); 52(12)7211–7224,
https://doi.org/10.1093/nar/gkae311.
4.
Xie,
J., Wang, L. & Lin, RJ. Variations of
intronic branchpoint motif: identification and
functional implications in splicing and disease.
Commun. Biol. 6, 1142 (2023). https://doi.org/10.1038/s42003-023-05513-7. 5.
Xie
J and Friedman R, Editorial: Evolution in
Neurogenomics. Front. Genet. (2023)14:1220750.
doi:
10.3389/fgene.2023.1220750. 6.
Liu
L, Das U, Ogunsola S, Xie J. Transcriptome-Wide
Detection of Intron/Exon Definition in the
Endogenous Pre-mRNA Transcripts of Mammalian
Cells and its Regulation by Depolarization, Int.
J. Mol. Sci. (2022) 23, 10157. https://
doi.org/10.3390/ijms231710157 7.
Tian
L, Xie X, Das U, Chen Y, Sun Y, Liu F, Lu H,
Peng N, Zhu Y, Gu X, Deng H, Xie J, Zhao X.
Forming cytoplasmic stress granules PURα
suppresses mRNA translation initiation of IGFBP3
to promote esophageal squamous cell carcinoma
progression, Oncogene, 41,
4336–4348 (2022), https://www.nature.com/articles/s41388-022-02426-3. 8.
Ling
Liu, Jinghua Feng, Julian Polimeni, Manli Zhang,
Hai Nguyen, Urmi Das, Xu Zhang, Harminder Singh,
Xiao-Jian Yao, Etienne Leygue, Sam K.P. Kung,
and Xie J. Characterization of cell free plasma
methyl-DNA from xenografted tumours to guide the
selection of diagnostic markers for early-stage
cancers. Frontiers in Oncology,
11:615821 (2021) Feb. 5. DOI: 10.3389/fonc.2021.615821. 9.
Xie
J, Weiskirchen R. What does the 'AKT' stand
for in the name 'AKT kinase'?: some historical
comments. Frontiers in Oncology,
10:1329. June, 2020, https://www.frontiersin.org/articles/10.3389/fonc.2020.01329/full. 10.
Nguyen H, Das U, Xie J. Genome-wide
evolution of wobble base-pairing nucleotides of
branchpoint motifs with increasing organismal
complexity. RNA Biology,
17:3, 311-324, Dec. 2019, http://dx.doi.org/10.1080/15476286.2019.1697548. 11.
Nguyen H, Xie J. Widespread separation of
the polypyrimidine tract from 3' AG by G tracts
in association with alternative exons in metazoa
and plants. Frontiers in Genetics,
9:741, published: 14 January 2019, doi:
10.3389/fgene.2018.00741. |PDF| 12.
Das U, Nguyen H, Xie J. Transcriptome
protection by the expanded family of
hnRNPs. RNA Biology, 16:2,
155-159, 2018 Dec 30. PMID: 30596342 DOI:
10.1080/15476286.2018.1564617. 13.
Lei L, Cao W, Liu L, Das U, Wu Y, Liu G, Sohail
M, Chen Y, Xie J. Multi-level differential
control of hormone gene expression programs by
hnRNP L and LL in pituitary cells. Mol.
Cell. Biol., 2018 May
29;38(12):e00651-17.(MCB Most-Read'04-05'18).
|PDF|
14. Nguyen H, Das U,
Wang B, Xie J. The matrices and
constraints of GT/AG splice sites of more
than 1000 species/lineages. Gene,
2018, 660:92-101.
|
|
|
The REPAG element specifically
among mammals |
|
||
|
The CaRRE element among
vertebrates (Liu GD, et al., J. Biol.
Chem. 2012, 287:22709–22716) |
|
|
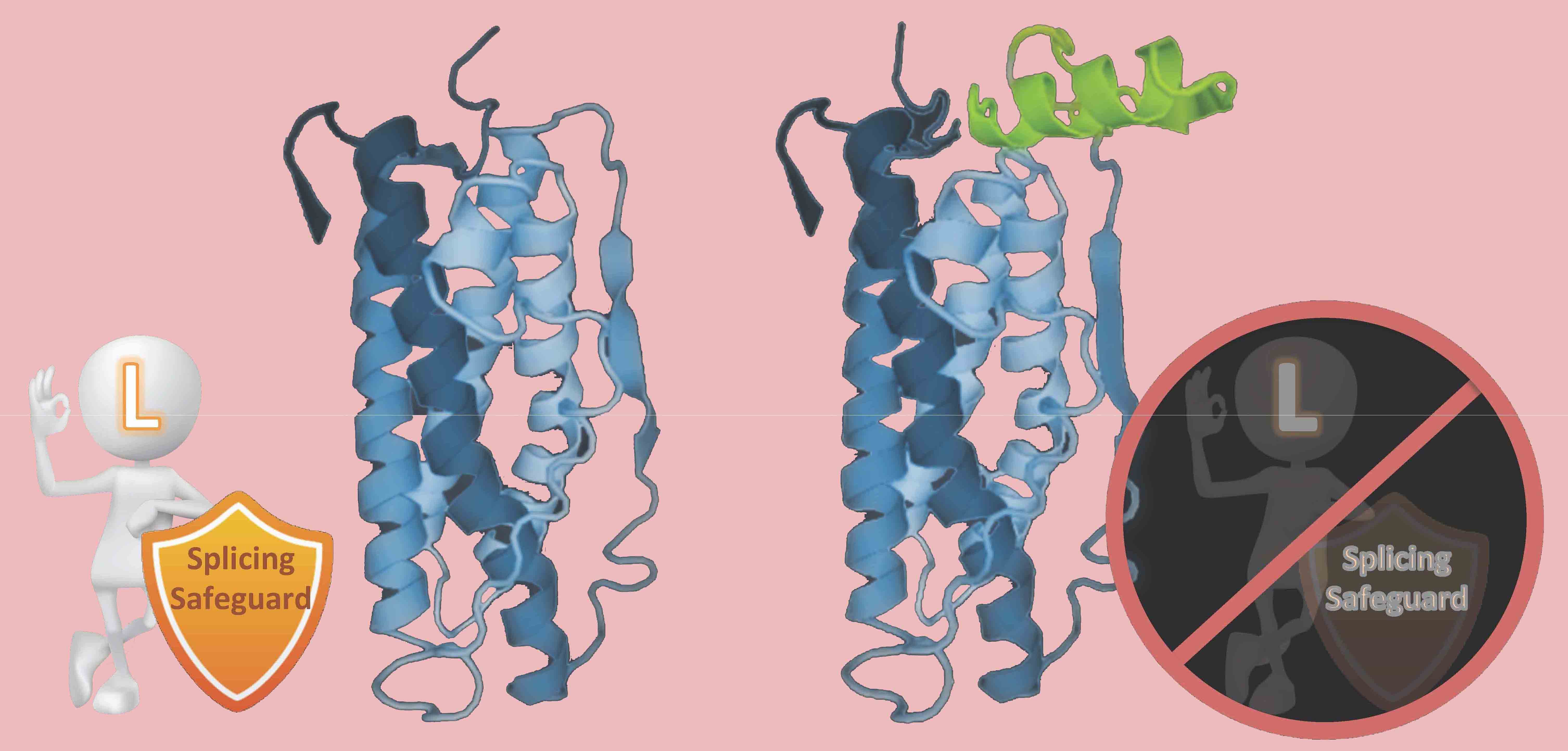
A new group of
introns: REPA element 'inserted' between the
Py and 3'AG and its effects on alternative
splicing |
|
*Research
in this lab has been supported by CIHR , NCIC, CBCF,
NSERC, MMSF, MHRC and CFI.
Misce. links:
- Eradication of Polio in China
 RNA Meeting in
Winnipeg, June 23-26, 2019 Some RNA/DNA
literature/history:
RNA Meeting in
Winnipeg, June 23-26, 2019 Some RNA/DNA
literature/history:- Why are the snRNAs in splicing called
U1, U2, U4, U5, U6 snRNAs? Here is the original paper
that describes the use of Us in the names for some of
the small nuclear RNAs that are Uridylic acid-rich, by
James L. Hodnett and Harris Busch of the Baylor
University College of Medicine in 1968: Hodnett JL,
Busch H. Isolation and characterization of uridylic
acid-rich 7 S ribonucleic acid of rat liver nuclei. J
Biol Chem. 1968 Dec 25;243(24):6334-42. |PDF|
- 125 Questions in
Science,
which ones are related to Alternative Splicing?
- How to present a clean, highlight
revision text without the deleted words in the
text or balloons after tracking changes in WORD?
- Albert Einstein the mediocre:
Why the h-index is a bogus measure of academic
impact (Reader discretion is advised).
- Sequencing RNA and its
modifications Webinar
- Opinions about the term non-coding RNA
- 谁翻译了基 因一词?维 基百科
Faculty of Graduate Studies |
Department of Physiology & Pathophysiology |
College of Medicine |
Faculty of Health Sciences |
University of Manitoba |